 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 8, 2010, pp. 949-956 REVIEW Overlooked post-translational modifications of proteins in Plasmodium falciparum: N- and O-glycosylation - A Review Cristiana S de MacedoI; Ralph T SchwarzII; Adriane R TodeschiniI; Jose O PreviatoI; Lucia Mendonça-PreviatoI, + IInstituto
de Biofísica Carlos Chagas Filho, Centro de Ciências da Saúde,
Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brasil Received 25 September
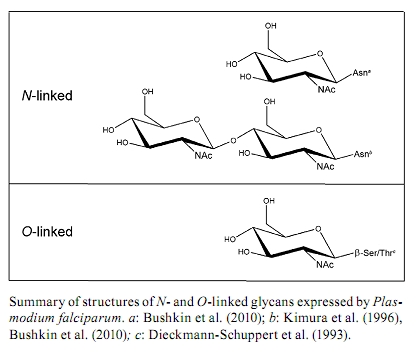
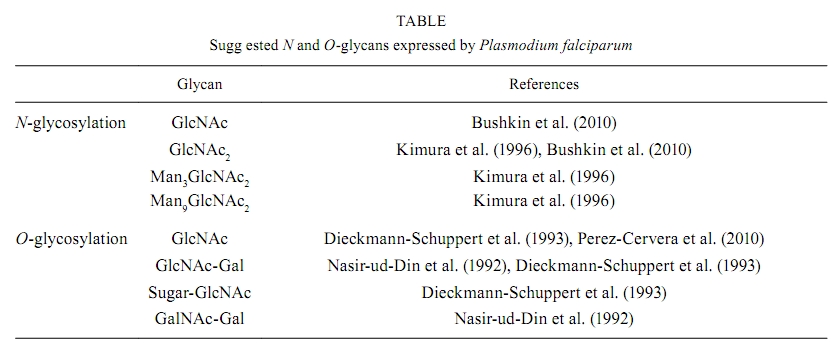
2010 Code number: oc10164 ABSTRACT Human malignant malaria is caused by Plasmodium falciparum and accounts for almost 900,000 deaths per year, the majority of which are children and pregnant women in developing countries. There has been significant effort to understand the biology of P. falciparum and its interactions with the host. However, these studies are hindered because several aspects of parasite biology remain controversial, such as N- and O-glycosylation. This review describes work that has been done to elucidate protein glycosylation in P. falciparum and it focuses on describing biochemical evidence for N- and O-glycosylation. Although there has been significant work in this field, these aspects of parasite biochemistry need to be explored further. Key words: glycoproteins - glycosylation - N-glycan - O-glycan - Plasmodium Malaria is a major human parasitic disease. According to the World Malaria Report (WHO 2008), 3.3 billion people live at risk for malaria infection. About 247 million episodes are reported every year, leading to 881,000 deaths, of which 85% are children under five years old. Malaria is caused by parasites from the Plasmodium genus. There are 156 different species that infect vertebrates, four of which infect humans: Plasmodium falciparum, Plasmodium vivax, Plasmodium ovale and Plasmodium malariae (Warrell & Gilles 2002). Recently, Plasmodium knowlesi, which causes malaria in monkeys, was detected in many cases of human malaria in Asia using molecular techniques (Singh et al. 2004). P. vivax, P. ovale and P. malariae cause infections that are seldom deadly, known as benign tertian malaria (P. vivax and P. ovale) and quartan malaria (P. malariae). These names refer to the periodicity of the fever attacks, which occur at the end of the intraerythrocytic cycle when infected erythrocytes break up (every 48 h in the case of P. vivax and P. ovale or every 72 h for P. malariae) and this event releases merozoites and stimulates the secretion of inflammatory cytokines by the host (Clark et al. 2006). P. falciparum causes infections that can lead to death if they are not treated appropriately. Because it has an intraerythrocytic cycle of 48 h and thus results in fever every three days, P. falciparum infection is known as malignant tertian malaria. Malaria caused by P. knowlesi infection is fatal in about one in 10 patients (Daneshvar et al. 2009). The life cycle of Plasmodium parasites alternates between a definitive host (a mosquito from the genus Anopheles), in which sexual reproduction takes plac, and an intermediary host (a vertebrate), in which parasite reproduction is asexual. Plasmodium species that infect humans (P. falciparum, P. vivax, P. ovale, P. malariae and P. knowlesi) have similar life cycles with slight variations. The Plasmodium life cycle begins in humans after an infected Anopheles female draws blood from the human during the blood meal and the mosquito injects saliva with sporozoites into the skin (Sinnis & Zavala 2008). A few hours after infection, the sporozoites reach the liver by traversing the Kupffer cells, where they begin to invade hepatocytes (Vaughan et al. 2008, Ejigiri & Sinnis 2009). This phase is known as the first asexual reproduction phase, or exoerythrocytic schizogony. This phase of the cycle varies between species: P. falciparum and P. malariae begin asexual replication immediately while P. vivax and P. ovale experience a dormant period inside hepatocytes. This latent stage is called the hypnozoite stage (Krotoski et al. 1982) and this stage can persist for long periods of time before the parasites undergo asexual replication to cause a primary infection. Hypnozoites are responsible for malaria relapses (Krotoski 1989) and they have not been detected in P. knowlesi infections in monkeys (Krotoski & Collins 1982). The asexual hepatic schizogony is a phase of significant parasite growth and division, during which hepatic schizonts are observed. These hepatic schizonts contain millions of merozoites, which are released into the blood stream and invade erythrocytes. These merozoites begin the intraerythrocytic schizogony phase of the cycle, during which symptoms of the malaria infection are presented. Giemsa staining has been used to detect infected erythrocytes in thick and thin blood smears of infected patients and blood smear microscopy is still the standard method for malaria diagnosis (Ndao 2009). After invasion, the merozoite undergoes differentiation into early trophozoite (0-20 h), known as the ring stage due to its morphology. Within 20-40 h, the mature trophozoite develops and exhibits an increased metabolism, during which erythrocyte cytoplasm is ingested and haemoglobin is digested. The toxic free haem moieties that are released are inactivated by the parasite through polymerization, giving rise to hemozoin, also called malarial pigment (Pisciotta & Sullivan 2008). After these events, nuclear division takes place to give rise to the schizont. Following maturation (40-48 h), schizonts rupture leading to the release merozoites into the blood stream. Newly released merozoites will either invade other erythrocytes, thereby beginning a new cycle of infection (Bannister & Mitchell 2009), or they will differentiate into sexual forms called gametocytes, which are either macrogametocytes (female) or microgametocytes (male) (Alano 2007). In the case of P. falciparum, merozoites appear 7-15 days after the onset of the intraerythrocytic cycle and they may circulate in the blood anywhere from 2 days-4 weeks (Talman et al. 2004). The Anopheles mosquito ingests circulating gametocytes with the blood meal. Inside the mosquito, gametocytes undergo a temperature drop (from 37-32ºC) as well as pH changes (from 7.4-8.2). These factors, together with exposure to xanthurenic acid, a gametocyte-activating factor present in the mosquito (Billiker et al. 1998, Hirai et al. 2001, Kuehn & Pradel 2010), induce gametogenesis, the sexual phase of the Plasmodium life cycle. The male and female gametes are formed during this stage: the microgamete (male) is formed through a process known as exflagellation and it fertilises the macrogamete (female) to form a zygote (Alano 2007). After zygote development, the resulting ookinete will migrate through gut epithelial cells (Baton & Ranford-Cartwright 2005) where it forms an oocyst, which then undergoes multiple asexual replications to form sporozoites. The sporozoites are released by oocyst rupture into the body cavity of the mosquito and migrate to and infect the salivary glands (Matuschewski 2006). Thus, sporozoites are able to invade two different target cell types: liver cells in the vertebrate host and salivary gland cells in the mosquito vector. When the infected mosquito bites another vertebrate host, the infection is spread. P. falciparum remains the best studied Plasmodium species due to the establishment of adequate culture methods (Trager & Jensen 1976). Although genome sequencing (Gardner et al. 2002) together with investigations of the biochemistry, molecular biology, cell biology and host-parasite interactions have been published for P. falciparum, protein glycosylation remains controversial because there is little information about the function of P. falciparum glycosylated molecules and because the few studies that are available are inconsistent due to differing methodologies. This review mainly focuses on N- and O-glycosylation in P. falciparum because glycosylphosphatidylinositol (GPI) anchors are already well established in this parasite (Gerold et al. 1994, 1996a, Hoessli et al. 1996, Davidson & Gowda 2001, Guha-Niyogi et al. 2001, Mendonça-Previato et al. 2005, von Itzstein et al. 2008, Chung et al. 2009, Shams-Eldin et al. 2009a, b, Debierre-Grockiego & Schwarz 2010). Glycosylated proteins in P. falciparum The first studies on glycosylation of Plasmodium proteins started with work by Trigg et al. (1977). These authors reported labelling of P. knowlesi-infected erythrocytes with galactose oxidase and tritiated sodium borohydride, which specifically label terminal galactose and galactosamine residues in glycolipids and glycoproteins (Gahmberg & Hakomori 1973). Infected erythrocytes exhibited lower levels of labelled glycoproteins than uninfected erythrocytes, possibly as a result of infection-association changes in the host-derived components of the erythrocyte membrane. Galactose oxidase binds to terminal and penultimate galactose residues in sialoglycoproteins and since Plasmodium organisms do not synthesise sialic acids (Schauer et al. 1984), they probably lack the structures that are typically labelled by this method. The presence of glycoproteins in P. falciparum has been demonstrated in several studies. Kilejian and Olson (1979) demonstrated the presence of P. falciparum proteins that were able to bind concanavalin A (Con A), which suggested the presence of mannosylated glycans. Later, Kilejian (1980) demonstrated the presence of glycoproteins, mainly in the late trophozoite and schizont stages, by radioactive labelling with glucosamine. Glycoproteins were also observed in merozoites in this study. There were 12 labelled proteins observed in the schizont stage; of those proteins, five remained in the merozoites and one was identified on the infected erythrocyte surface. Udeinya and Van Dyke (1980) treated P. falciparum-infected erythrocytes with radioactive sugars (glucosamine, fucose and mannose) to identify glycoproteins in these samples. In agreement with Kilejian (1980), the incorporation of radioactive glucosamine was stronger for erythrocytes infected with parasites in later stages (trophozoites and schizonts). A more detailed study identified eight labelled glycoproteins, ranging from 19-90 kDa, that were incorporated into the membrane (Udeinya & Van Dyke 1981a). These results might reflect the biosynthesis of GPI anchors, which are known to be upregulated in trophozoites and schizonts (Schmidt et al. 1998). In the same year, David et al. (1981) demonstrated that erythrocytes infected with later stages of P. falciparum preferentially bound to agarose beads coated with lectins (Con A, Ricinus communis lectin, soybean lectin and peanut agglutinin), which was in contrast with ring-infected and uninfected erythrocytes. These observations suggested that P. falciparum infection might result in increased mannosyl and/or glucosyl residues as well as increased galactosyl residues on the erythrocyte membrane. This observation could be a result of glycoprotein production by the parasite or infection-induced modifications of erythrocyte proteins. Ramasamy (1987) later confirmed the presence of P. falciparum glycoproteins that could bind to Con A, wheat germ agglutinin (WGA), R. communis lectin 120 and Bandeirea simplicifolia lectin (BS). Binding to WGA exhibited the highest affinity, thereby suggesting the presence of N-acetyl glucosamine (GlcNAc) residues. Perkins (1982) investigated the incorporation of radioactive methionine, glucosamine and mannose into the surface of schizont-infected erythrocytes as well as in the merozoite stage by adding proteolytic (trypsin and pronase) and glycolytic (neuraminidase) enzymes. In this study, glycoproteins were released from schizont-infected cells upon pronase treatment but not after neuraminidase treatment, suggesting the absence of sialic acids in P. falciparum. In contrast to the conclusions of Kilejian (1980), this study did not identify glycoproteins in merozoites. Further studies demonstrated the importance of glycosylated proteins in the immunogenicity of P. falciparum. Ramasamy and Reese (1985) observed that treating P. falciparum antigens (obtained from schizont-infected erythrocytes) with mixed glycosidases from Charonia lampas (sea snail) decreased their affinity to antibodies from infected hosts, such as owl monkeys that were immunised with P. falciparum and Nigerian subjects living in malaria endemic areas. Specifically, two parasite proteins at 65 and 100 kDa exhibited decreased reactivity after enzyme treatment, and the carbohydrate modification of the epitopes was confirmed by their insensitivity to protease activity and their heat stability. Endoglycosidase F (PNGase F) treatment had little effect on the reactivity of the antibodies, which suggested that N-linked glycans exhibit little contribution to this effect. A more detailed study (Ramasamy & Reese 1986) demonstrated that antibody binding was abolished by treatment with α-galactosidase obtained from Aspergillus niger and green coffee beans. A mixture of α-galactosidase, fucosidase, mannosidase and N-acetyl hexosaminidase also abolished antibody binding. Several antigens were not recognised by the antibodies after treatment with these enzymes, particularly, antigens at 185, 135, 120 and 75 kDa. Notably, the enzyme mixture was ineffective without α-galactosidase, suggesting an important role for terminal α-galactosyl residues in the antigenicity of P. falciparum glycoproteins. Supporting these results, a fluorescent derivative of BS that specifically binds to α-galactosidase demonstrated binding to acetone-fixed parasites (Jakobsen et al. 1987). In addition, antibody reactivity to soluble P. falciparum molecules was sensitive to α-galactosidase treatment. Later, Ravindran et al. (1988) reported that sera from subjects living in endemic areas, as well as from patients with acute P. falciparum infection, exhibited high titres of antibodies specific for α-galactose, which was in contrast to sera from patients with acute cerebral malaria. Panton et al. (1989) demonstrated that the histidine-rich protein II (HRP-II) from P. falciparum, an extracellular protein found in sera from infected patients and in the supernatant of cultured parasites, could be labelled with radioactive galactose. However, intracellular HRP-II was not labelled, suggesting that glycosylation might act as a signal for secretion. In 1990, Jackobsen et al. described a soluble 60 kDa glycoprotein that was present on the surface of trophozoites and schizonts, but not merozoites, and this protein was able to bind to Erythrina christagalli agglutinin, but not to Con A or Griffonia simplicifolia agglutinin, suggesting the presence of β-D-galactose(1-4)-D-N-acetylglucosamine. This protein was recognised by human sera from different endemic regions, as well as by lymphocytes from malaria-immune individuals. Other studies on P. falciparum immunogenic proteins, specifically the merozoite surface protein (MSP), demonstrated that both MSP-1 (195 kDa) and MSP-2 (43 kDa) could be galactosylated after metabolic labelling with tritiated N-acetylglucosamine, and both proteins were digested with α and β-galactosidases. However, the incorporation of galactose into the GPI moiety of this protein could not be ruled out (Qazi et al. 1994). Furthermore, Klabunde et al. (2002) identified P. falciparum glycoproteins that bound to mannan-binding protein during innate immunity. The nature and structure of the glycans were not specifically addressed in these early studies, but later studies verified the presence of N- and O-glycosylation in P. falciparum. N-glycosylation in P. falciparum Early studies of N-glycosylation in P. falciparum made use of inhibitors, namely, tunicamycin and 2-deoxyglucose (Udeinya & Van Dyke 1980, 1981b, c). Both compounds arrested parasite development in a dose-dependent manner and both inhibited the incorporation of radiolabeled glucosamine into glycolipids, but not the incorporation of isoleucine into proteins. These findings suggested the specific inhibition of N-glycosylation, but the incorporation of glucosamine into O-glycans or GPI anchors was not considered. Tunicamycin blocks N-glycosylation by inhibiting UDP-GlcNAc:dolichol-pyrophosphate N-acetylglucosaminyltransferase (Schwarz & Datema 1982, McDowell & Schwarz 1988). However, tunicamycin also blocks ganglioside biosynthesis in the Golgi apparatus of rat liver (Yusuf et al. 1983a, b). Alternatively, 2-deoxyglucose blocks N-glycosylation through the formation of dolichyl-PP-(GlcNAc)2-2-deoxyglucose, thereby preventing the elongation of the mannose chain (Schwarz & Datema 1982, Elbein 1987, McDowell & Schwarz 1988). Notably, 2-deoxyglucose exhibits broader effects on P. falciparum metabolism and it blocks both glycolysis (van Schalkwyk et al. 2008) and GPI biosynthesis (Udeinya & Van Dyke 1981c, Dieckmann-Schuppert et al. 1992a, Santos de Macedo et al. 2001, de Macedo et al. 2003). Therefore, the arrest in parasite development that was observed after tunicamycin or 2-deoxyglucose treatment cannot be completely attributed to the inhibition of N-glycosylation. Dieckmann-Schuppert et al. (1992a, b) reported that tunicamycin could not inhibit P. falciparum growth in the first life cycle (48 h). However, when parasites were subsequently treated with tunicamycin at concentrations that would not affect protein biosynthesis (12 μM), parasites underwent arrested development, resulting in death. These observations were confirmed by Kimura et al. (1996). Furthermore, Dieckmann-Schuppert et al. (1992b, 1996) reported that lower concentrations of tunicamycin reduced the incorporation of radioactive methionine by 30% after 30 min of treatment, which was not observed by Kimura et al. (1996). Therefore, the exact biochemical effects of tunicamycin on P. falciparum metabolism remain unclear. The first attempts to study N-glycosylation in P. falciparum protein investigated the sensitivity of the parasite proteins to peptide N-glycosidase F (also known as PNGase F and N-glycanase®), but these initial studies gave negative results (Ramasamy 1987, Dayal-Drager et al. 1991). Using metabolic labelling, Dieckmann-Schuppert et al. (1992a) demonstrated that PNGase-F treatment led to the release of a small percentage of tritiated glucosamine (7-12%) from proteins. While this finding supported the presence of some protein N-glycosylation, it was considered to be negligible at the time. Furthermore, dolichol-PP-linked intermediates, which would be indicative of dolichol-cycle intermediates, were not found in this study. In addition, P. falciparum lysates did not catalyse the transfer of radiolabeled sugars from dolichyl-pyrophosphoryl-oligosaccharides [Dol-PP-(GlcNAc)2, Dol-PP-(GlcNAc)2-Man9 or Dol-PP-(GlcNAc)2Man9Glc] to peptide acceptors of N-glycosylation. In contrast, after radiolabeling with radioactive methionine for 18 h, Kimura et al. (1996) identified proteins that bound to Con A-Sepharose that were absent when parasites were pretreated with tunicamycin for 52 h before labelling. The same proteins lost their affinity to Con A-Sepharose after PNGase F treatment, which indicates that these proteins were N-glycosylated. Glycans released from these parasites were analysed by thin layer chromatography to identify oligosaccharides that migrated between GlcNAc2, Man3GlcNAc2 and Man9 GlcNAc2 standards. Because tunicamycin inhibited the transition of late trophozoites into schizonts and because N-glycosylated proteins were not observed in parasites that exhibited arrested development, these N-glycosylated proteins could play a role in the development of the intraerythrocytic stages of P. falciparum. The inconsistent results obtained by Dieckmann-Schuppert et al. (1992a) and Kimura et al. (1996) could be attributed to different experimental approaches. While Dieckmann-Schuppert et al. (1992a) analysed late trophozoites, Kimura et al. (1996) used asynchronous cultures that contained three different stages (ring, trophozoite and schizont) after Percoll® gradient purification. In addition, Dieckmann-Schuppert et al. (1992a) used shorter pulse times (4 h) and glucose-free medium in their radiolabeling procedure. In contrast, Kimura et al. (1996) radiolabelled for both 8 h and 18 h and used glucose and methionine-deficient media because glucose-free medium had been reported to interfere with N-glycan biosynthesis in other systems (Elbein 1987). Gowda et al. (1997) confirmed the observations of Kimura et al. (1996) using similar radiolabeling conditions and they claimed to be the first to rule out GPI biosynthesis as an alternative explanation for the common experimental observations. A later study (Schmidt et al. 1998) demonstrated that GPI biosynthesis increases by 21-fold during intraerythrocytic development from the ring stages (6 h) to the late schizont stages (46 h). These data suggests that GPI biosynthesis is incipient during stages that also exhibit N-glycosylation, thereby supporting earlier conclusions from Kimura et al. (1996). N-glycosylation was also analysed for other P. falciparum proteins, such as MSP-1 and MSP-2. Neither MSP-1 nor MSP-2 exhibited lectin affinity or sensitivity to PNGase F, suggesting absence of N-linked glycans despite the presence of several potential N-glycosylation sites in both proteins (Berhe et al. 2000). Notably, when MSP-1 was expressed in mammalian cells (Murphy et al. 1990, Burghaus et al. 1999, Yang et al. 1999) and in the baculovirus system (Kedees et al. 2000), the proteins do become N-glycosylated. The availability of the P. falciparum genome (Gardner et al. 2002) allowed for new studies on enzymes involved in N-glycosylation. P. falciparum lacks most of the enzymes involved in the assembly of N-glycans and phylogeny analyses suggested a secondary loss of Alg glycosyltransferases. The only two enzymes predicted to be present in P. falciparum were Alg7 and Alg14, which are responsible for the synthesis of Dol-PP-GlcNAc and Dol-PP-GlcNAc2, respectively, and STT3, which is responsible for the transfer of the Dol-PP-oligosaccharide to the Asn residue in the N-X-T/S sequon (Templeton et al. 2004, Samuelson et al. 2005, Anantharaman et al. 2007). Further biochemical studies confirmed that P. falciparum synthesises N-glycans that are shorter than their mammalian counterparts, namely, Dol-PP-GlcNAc and Dol-PP-GlcNAc2. The type of N-glycan that is produced is stage-dependent: early ring stages synthesise mainly Dol-PP-GlcNAc whereas schizonts synthesise both Dol-PP-GlcNAc and Dol-PP-GlcNAc2. These glycans are transferred to proteins, such as glucosamine-labelled parasite proteins, and they are recognised by the GlcNAc-binding G. simplicifolia lectin (GSL-II). This recognition is attenuated by PNGase F treatment, which decreases lectin binding (Bushkin et al. 2010). Although Kimura et al. (1996) did detect GlcNAc and GlcNAc2 among the glycans released by PNGase F treatment, their detection of larger glycans that were considered to be mannosylated is not fully understood. The N-glycan biosynthetic limitations exhibited by Plasmodium could explain why Dieckmann-Schuppert et al. (1992a, 1994) did not observe the transfer of radiolabeled Dol-PP-(GlcNAc)2Man9 or Dol-PP-(GlcNAc)2Man9Glc to acceptors in a cell-free system because, in this system, the oligosaccharyltransferase would not recognise these structures. However, when exogenous peptide acceptors were used, glycosylation by endogenous Dol-PP-GlcNAc or Dol-PP-GlcNAc2 was also not observed (Dieckmann-Schuppert et al. 1992a, 1994), which may have resulted from the limited size of the internal pool of Dol-PP-GlcNAc and/or Dol-PP-GlcNAc2, thus making the detection of the corresponding reaction products difficult. Bushkin et al. (2010) demonstrated that fluorescent GSL-II stained the endoplasmic reticulum, rhoptries and the parasite surface, but it did not stain the apicoplast, food vacuole, parasitophorous vacuole or the infected erythrocyte. This staining pattern may have resulted from the inability of GSL-II to label proteins decorated only with GlcNAc. In any case, GSL-II labelling of some but not all Plasmodium organelles suggests that N-glycosylation may have a role in protein targeting, which will need to be analysed in more detail in future experiments (Figure, Table). O-glycosylation in P. falciparum Dayal-Drager et al. (1991) investigated the nature of the sugar linkage to proteins in P. falciparum. Endo-O-glucosidase (O-glycanase®) released sugars from parasite proteins labelled with tritiated mannose and glucosamine and the presence of hexosamines linked to serine residues was confirmed by β-elimination/reduction. Nasir-ud-Din et al. (1992) obtained similar results for a 195 kDa merozoite glycoprotein, probably MSP-1, thereby confirming the presence of O-GlcNAc, O-GalNAc and galactose in this protein. Dieckmann-Schuppert et al. (1992a) treated galactose, fucose and glucosamine-radiolabeled proteins with PNGase-F to remove possible N-glycans, purified the resulting material by gel filtration and subjected the void volume fraction to alkaline β-elimination. About half of the radioactivity was shifted to the monosaccharide volume, suggesting the release of O-glycans. Dieckmann-Schuppert et al. (1993) characterised O-glycosylation reactions using exogenous galactosylation to identify O-GlcNAc in proteins. An analysis of the alditols released after β-elimination demonstrated the presence of both reducing terminal and nonterminal GlcNAc, which could be elongated. O-GlcNAc transferase activity in parasite lysates was confirmed using exogenous peptide acceptors with a Pro-Tyr-Thr-Val-Val sequence and activity was higher in the later parasite stages. In contrast to the results of Nasir-ud-Din et al. (1992), mucin-type O-glycans were not detected in this study, which is consistent with later findings reporting that this parasite does not possess GalNAc-decorated proteins and is unable to synthesise GalNAc. Radiolabeling with GalN demonstrated that this precursor was mostly phosphorylated while GalNAc, GlcNAc and the corresponding sugar nucleotides were not detectable. These results are consistent with previous data that suggested that Plasmodium organisms are unable to epimerise GalN into GlcN and vice-versa, as shown by the absence of detectable GalN after labelling with GlcN (Dieckmann-Schuppert et al. 1992a, 1993, 1996). Kimura et al. (1996) confirmed the presence of O-linked glycoproteins that were mostly present in the schizont stage, which is in agreement with the identification of an O-GlcNAc modified protein in the same stage (Dieckmann-Schuppert et al. 1993). However, these authors suggested that the sensitivity to O-glycanase® could be attributed to the presence of Gal-GalNAc, which is in contrast to the observations of Dieckmann-Schuppert et al. (1992a, 1993). In contrast, Gowda et al. (1997) suggested that O-glycosylation does not take place in P. falciparum intraerythrocytic stages. Furthermore, in silico studies on O-glycosylation in P. falciparum suggested that this species lacks the genes that are necessary for the synthesis of mucin-type O-glycans (Templeton et al. 2004), which supports the findings from Dieckmann-Schuppert et al. (1992a, 1993). The presence of O-glycosylation in MSP from P. falciparum remains controversial. Nasir-ud-Din et al. (1992) observed that MSP-1 was modified by O-GlcNAc. Later, Khan et al. (1997) concluded that MSP-2 was O-glycosylated using asynchronous P. falciparum cultures. These authors employed β-elimination, exogenous galactosylation assays, lectin binding assays and enzymatic treatments to investigate O-glycosylation, and they observed that GlcNAcβ-Ser/Thr was the predominant structure whereas Gal-GalNAcα-Ser/Thr was also present to a minor degree. This second conclusion is incompatible with Dieckmann-Schuppert et al. (1992a, 1993), which reported the absence of GalNAc metabolism in P. falciparum. In contrast to the findings of Khan et al. (1997), Berhe et al. (2000) used a similar experimental approach to show that MSP-1 and MSP-2 obtained from merozoites were not modified with O-glycans because β-elimination caused no effect on the migration of glycopeptides in a Bio-Gel P4 column. These conclusions were further supported using galactosylation and lectin binding assays. Neither MSP-1 nor MSP-2 incorporated labelled galactose or bound to the lectins in the assay, indicating that the sole post-translational modification of this protein was the GPI anchor (Gerold et al. 1996b). Hoessli et al. (2003) showed by immunoprecipitation that, in addition to the 19-kDa C-terminal fragment that contains the GPI anchor, MSP-1 fragments could be labelled with tritiated glucosamine. In addition, they also demonstrated that both N and C-terminal MSP-1 fragments, as well as the whole protein, could be exogalactosylated. Furthermore, five different MSP-1 sequences from various P. falciparum strains contained β-GlcNAc (but not α-GlcNAc) sites that were predicted in silico. Regarding nucleocytosolic O-glycosylation, which is the addition of O-GlcNAc to Ser/Thr residues, no O-linked N-acetylglucosaminyltransferase (OGT) related to other organisms was identified in the Plasmodium genome (Banerjee et al. 2009). However, recently, the presence of OGT has been investigated using antibodies against OGT, and these studies have suggested the presence of O-GlcNAcylation in P. falciparum (Perez-Cervera et al. 2010), supporting previous observations of GlcNAcβ-Ser/Thr in MSP-2 (Khan et al. 1997). Despite the progress made in elucidating the biology of malaria parasites and in the discovery of effective drugs and anti-vector measures, malaria still remains one of the most important diseases in the world. Treatment has become difficult due to the emergence of parasites that are resistant to chloroquine and pyrimethamine. The need for new antimalarial drugs has encouraged research to investigate the basic biochemistry of the malaria parasites. In the light of the results summarised in this review, it is clear that both N- and O-glycosylation occur in P. falciparum. However, the identity of the proteins that carry these modifications, as well as their biological significance in the parasite life cycle and their interactions with its vertebrate and invertebrate hosts remains to be elucidated. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10164t1.jpg] [oc10164f1.jpg] |
| |||||||||

{kind=link}
{kind=link}