 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
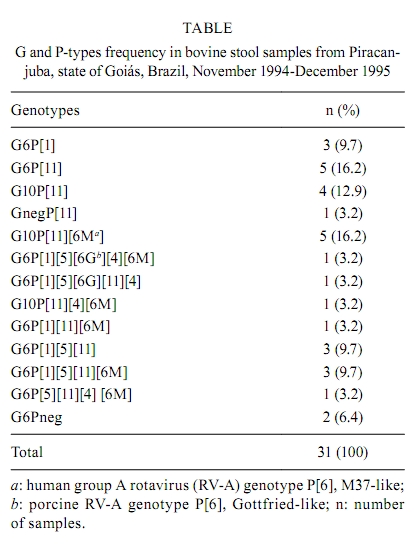
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 8, 2010, pp. 1040-1043 ARTICLES Molecular characterization of G and P-types bovine rotavirus strains from Goiás, Brazil: high frequency of mixed P-type infections Thabata Alessandra Ramos CaruzoI, +; Willia Marta Elsner Diederichsen de BritoII; Veridiana MunfordIII; Maria Lúcia RáczIII IDepartamento
de Genética, Evolução e Bioagentes, Instituto de Biologia,
Universidade Estadual de Campinas, Campinas, SP, Brasil Received 2 August
2010 Code number: oc10177 ABSTRACT In this study, 331 samples from calves less than one month old from a dairy herd in the district of Piracanjuba, state of Goiás, Brazil were tested for rotavirus. Thirty-three samples (9.9%) tested positive for rotavirus. Out of those, 31 were submitted to G and P characterization by reverse transcription followed by semi-nested polymerase chain reaction. Two samples were characterized as G6P[1], three as G10P[11] and five as G6P[11]. The majority of the samples (51.6%) displayed multiple P genotypes (P-genotype mixtures), including typical human genotypes P[4] and P[6M], suggesting the occurrence of co-infections and genetic reassortment. Also, the detection of human genotypes in bovine samples may be considered evidence of the zoonotic potential of rotaviruses. To our knowledge, this is the first report of such a high frequency of P genotype mixtures in bovine rotavirus samples. It also increases data on G and P rotavirus genotypes circulating in dairy herds in Brazil and can help in the development of more efficient immunization approaches, thereby controlling infection and reducing economical losses. Key words: bovine rotavirus - genotyping - mixed infections - reassortment - Brazil Rotaviruses have been recognized as important agents of acute diarrhoea in both humans and animals (Greenberg & Estes 2009) and are responsible for considerable economical losses worldwide. Members of the Reoviridae family, genus Rotavirus, rotaviruses are classified into seven groups, A-G (Estes & Kapikian 2007). The virion has a triple-layered protein capsid of 100 nm in diameter, surrounding a genome composed of 11 segments of double stranded RNA (dsRNA). Each segment encodes six structural and six non-structural proteins (Greenberg & Estes 2009). Group A rotaviruses (RV-A) are classified as G and P-genotypes according to two type-specific outer capsid proteins, VP7 (glycoprotein) and VP4 (protease sensitive), respectively. Genotype-specific classification is based on results from reverse transcription followed by polymerase chain reaction (RT-PCR) and nucleotide sequence analysis. Based on molecular differences, at least 23 different G-types and 32 P-types have been described so far ( Hoshino & Kapikian 2000, Rao et al. 2000, Martella et al. 2007, Ghosh et al. 2008, Greenberg & Estes 2009, László et al. 2009, Matthijnssens et al. 2009, Schumann et al. 2009, Solberg et al. 2009, Trojnar et al. 2009). Due to its segmented genome, genetic reassortment may occur when at least two different RV-A strains infect a single cell, resulting in (i) new strains, (ii) characterization of more than one G or P-type in one sample, (iii) introduction of animal rotavirus genes in human rotaviruses (HuRV) populations and vice-versa and (iv) natural RV-A reassortants infecting different animal species, as suggested by Dhama et al. (2009) in a recent report of the isolation of G3 and G5 bovine rotaviruses (BoRV). Also, because VP4 and VP7 genes independently segregate, many G and P combinations can be observed in natural infections (Iturriza-Gomara et al. 2001, Palombo 2002, Leite et al. 2008, Matthijnssens et al. 2008a, Dhama et al. 2009, Martella et al. 2010). BoRV-A have been classified as G6, G8 or G10 genotypes, associated with either P[1], P[5] and/or P[11] and genotypes G6P[5], G6P[1] and G10P[11] are considered the most common ones. However, genotypes G1, G3, G5, G7, G11, G15 and P[7], P[14], P[17] and P[21] have also been described for BoRV-A samples, but less frequently (Hussein et al. 1993, Fukai et al. 1999, Rao et al. 2000, Okada & Matsumoto 2002, Ghosh et al. 2008). SUBJECTS, MATERIALS AND METHODS A total of 331 stool samples from calves less than one month old from a dairy herd in the district of Piracanjuba, state of Goiás, Brazil was collected directly from the rectum during November and December of 1994 and 1995. RV-As were detected by enzyme immunoassay for rotavirus and adenovirus (EIARA-Fiocruz), in accordance with Pereira et al. (1985) and polyacrylamide gel electrophoresis (PAGE) as described by Pereira et al. (1983). Extraction of dsRNA from 20% (w/v) faecal suspensions was performed using Trizol® Reagent (Invitrogen), as described by the manufacturer, and stored at -20ºC. Molecular characterization of G and P-types by semi-nested multiplex RT-PCR was adapted from Gouvea et al. (1990, 1993, 1994a, b), Gentsch et al. (1992) and Das et al. (1994) and the products were detected after separation by electrophoresis in 1.5% agarose gel. Super Script™One Step RT-PCR with Platinum®Taq kit (Life Technologies Co) was used as a secondary approach to genotype some P-type negative samples. For negative G-type samples, Platinum®Taq DNA Polymerase (Life Technologies Co) was used. In both approaches, the reaction was modified to include 1 h RT at 42ºC followed by 40 cycles of amplification, as described by Das et al. (1994). Adapted cell culture strains of simian rotavirus SA11 (P[2]G3), BoRV UK (P[5]G6), B223 (P[11]G10) and NCDV (P[1]G6) and HuRV Wa (P[8]G1), DS-1 (P[4]G2) and ST3 (P[8]G4) were kindly supplied by Dr DR Snodgrass (Moredun Research Institute, Scotland) and used as positive controls. Water was used as a negative control in all reactions. RESULTS A total of 33 (9.9%) samples tested positive for RV-A by EIARA and PAGE. Two samples were not submitted to molecular characterization because of insufficient stool. From a total of 31 BoRV-A samples, only three could not be completely characterized according to their G or P specificities. For the others, G and P-types and many P-type mixtures, including typical HuRV-A P-types, were characterized (Table). Strains showing mixtures of genotypes were submitted to single primer semi-nested RT-PCR to confirm the occurrence of more than one genotype, previously detected by multiplex RT-PCR. This precaution was taken to avoid false positive results that could occur due to primer mismatches during multiplex RT-PCR. DISCUSSION Over the years, many epidemiological studies of BoRV-A have reported G6 and G10 as the most common G-types found in herds around the world. In this study, G6 was the most prevalent genotype, accounting for 64.5% of all 31 samples and G10 was detected in 32.2% of them (Table). Similar results were found in Italy (Monini et al. 2008), as the authors described 78.5% of samples as G6 and 9.9% as G10. Regarding P-types, results found on this study may be considered very peculiar when compared to P-types most frequently, reported P[11] and P[5]. Despite the fact that 32.2% of all samples were characterized as P[11] and 9.7% as P[1], P-types mixtures were common in this set of samples, accounting for 51.6% of it (Table). For comparison, Monini et al. (2008) found no P[1] in their set of samples and 65.1% of P[11], 25% of P[5] and only 2.5% of P-type mixtures. As reported before, the most common VP7/VP4 combinations for BoRV-A are G6P[5] (UK-like), G6P[1] (NCDV-like) and G10P[11] (B223-like) (Hussein et al. 1993, Fukai et al. 1999, Rao et al. 2000, Okada & Matsumoto 2002, Ghosh et al. 2008). In this study, no samples were characterized as G6P[5]. Nevertheless, Garaicoechea et al. (2006) characterized 150 (60%) BoRV-A collected from beef and dairy herds in Argentina as G6P[5]. More recently, this genotype was found in 38.5% (10/26) of BoRV-A in Australia, as described by Swiatek et al. (2009). Also, G6P[5] has been reported as the most prevalent BoRV-A genotype (40%) in samples collected from herds in the South, Southeast and Central Regions of Brazil between 1996-1999, as described by Alfieri et al. (2004). In this study, only seven samples displayed the other two typical BoRV-A genotypes: three (9.7%) samples were characterized as G6P[1] and four (12.9%) as G10P[11]. The frequency of these genotypes is similar to that reported by Alfieri et al. (2004), who also tested Brazilian samples (G6P[1], 12%, and G10P[11], 16%). However, these frequencies are much higher than those found in other studies: Reidy et al. (2006), in Ireland, characterized only 1.9% of their 54 samples as G6P[1] and 1.9% as G10P[11], and Garaicoechea et al. (2006) found no G6P[1] samples and only 4.4% G10P[11]. Five (16.2%) samples were characterized as G6P[11]. Reidy et al. (2006) and Garaicoechea et al. (2006) also described this genotype in their studies, but, again, in lower frequencies (7.4% and 4.4%, respectively). These authors also characterized some samples as G10P[5]. All these findings could be a result of co-infection and genetic reassortment and they reinforce the idea that segments displaying VP4 and VP7 genes independently segregate in nature (Iturriza-Gomara et al. 2001, Maunula & Von Bonsdorff 2002, Matthijnssens et al. 2008a, Greenberg & Estes 2009). A total of 16 samples (51.6%) displayed P-type mixtures (Table), including HuRV-A typical genotypes (P[4] and P[6M]). Many of these strains were also characterized as G6 or G10, thus bringing up once more the idea of genetic reassortment and independent gene segregation during virus replication. Reidy et al. (2006) and Swiatek et al. (2009) also described P genotype mixtures, G6P[5][11], in some samples, but again in lower frequencies (11% and 15.4%, respectively) when compared to those found in our study (Table). Also, Brito et al. (2000) characterized two G10P[5][11] strains (2/11) collected from two dairy herds experiencing a rotavirus outbreak in the Central Region of Brazil. To our knowledge, this high frequency of P genotype mixtures has not been yet described for BoRV-A and it reaffirms RV-A's reassortment potential among animal strains and also between human and animal strains. Characterization of typical human rotavirus P-types in bovine faecal samples is supported by Matthijnssens et al. (2008a), who, after sequencing all 11 segments of many RV-A strains, found a great constellation of genes with many different origins. Also, Palombo (2002) states that interspecies transmission of rotavirus is not a rare event and may occur frequently in nature. These 41.9% bovine samples with typical HuRV-A P-types may result in spreading an RV-A displaying new G and P occurrences and combinations from cattle to human. This event can become a public health problem, especially in developing countries, where many children can get infected in remote areas and at country sites (Coluchi et al. 2002, Bányai et al. 2004, 2009, Ramani et al. 2009) and among people who manage herds elsewhere. These new viruses may or may not be infectious at first, but they can circulate in the population and undergo other genetic modifications and subsequent adaptations that may be responsible for diarrhoeal disease among humans. In this case, rotaviruses can be considered a zoonosis, as reviewed by Cook et al. (2004) and Martella et al. (2010). The occurrence of P[6] Gottfried-like genotype in two samples confirms that cattle and piglet co-infection with typical BoRV-A and porcine rotaviruses-A can occur, as reported over the years (Rácz et al. 2000, Palombo 2002, Parra et al. 2008). Recently, Matthijnssens et al. (2008a) characterized all 11 rotavirus gene segments of many strains, including bovine KJ44 and KJ75. These strains were found to have porcine characteristics in all their gene segments, except for VP3 and VP4, which had bovine characteristics. Results described in this paper reinforce the idea that rotavirus interspecies transmission, co-infection and genetic reassortment frequently occur in nature and that reassortment could be the main mechanism responsible for evolution and diversity of rotaviruses, as already stated (Iturriza-Gomara et al. 2001, Maunula & Von Bonsdorff 2002, Ghosh et al. 2008, Matthijnssens et al. 2008a, b, Greenberg & Estes 2009). Nevertheless, the uncommon results found in this paper will be confirmed by gene sequencing. This paper also provides new data on G and P-types circulating in dairy herds in Brazil and suggests that constant improvement of herd management and immunization approaches may be crucial for controlling rotavirus infections and reducing economical losses. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10177t1.jpg] |
| |||||||||

{kind=link}