 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 105, No. 8, 2010, pp. 1068-1072 TECHNICAL NOTES Rotavirus A genotype G1P[8]: a novel method to distinguish wild-type strains from the Rotarix® vaccine strain Tatiana L Rose+; Marize P Miagostovich; José Paulo G Leite Laboratório
de Virologia Comparada e Ambiental, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil
4.365, 21040-360 Rio de Janeiro, RJ, Brasil Received 31 May
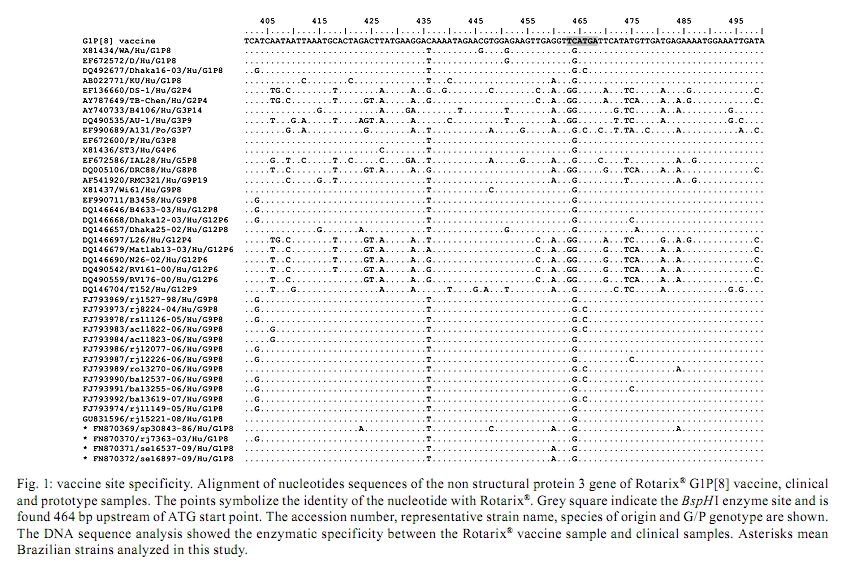
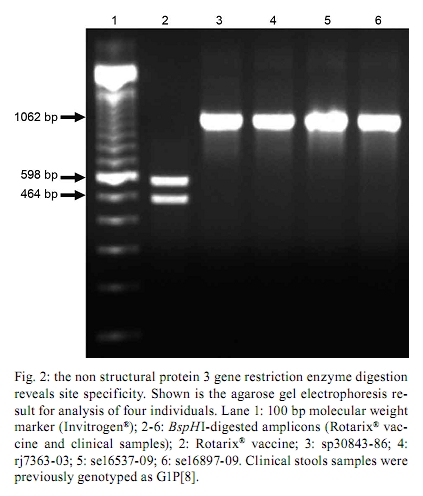
2010 Code number: oc10184 ABSTRACT Rotaviruses are important enteric pathogens for humans and animals. Group A rotaviruses (RV-A) are the most common agents of severe gastroenteritis in infants and young children and vaccination is the most effective method to reduce RV-A-associated diseases. G1P[8], the most prevalent RV-A genotype worldwide, is included in the RV-A vaccine Rotarix®. The discrimination between wild-type G1P[8] and vaccine G1P[8] strains is an important topic in the study of RV-A epidemiology to manage outbreaks and to define control measures for vaccinated children. In this study, we developed a novel method to segregate the wild-type and vaccine strains using restriction endonucleases. The dsRNA from the Rotarix® vaccine was sequenced and the NSP3 gene was selected as the target gene. The vaccine strain has a restriction pattern that is different than that of wild-type RV-A G1P[8] isolates after digestion with the restriction endonuclease BspHI. This pattern could be used as a marker for the differentiation of wild-type G1P[8] strains from the vaccine strain. Key words: Rotarix® vaccine strain - G1P[8] wild-type strain - rotavirus A strain differentiation - restriction endonuclease Group A rotavirus (RV-A) infection is a global public health concern and an important cause of paediatric hospitalization due to severe diarrhoea (Parashar et al. 2006, 2009). RV-A transmission occurs mostly through the faecal/oral route via direct or indirect contact with infected individuals. Because RV-A infects children in both developed and developing countries, factors such as malnutrition and poverty may not play an important role in RV-A infections (Schael et al. 2009). The introduction of rotavirus vaccines into the national immunization schedule for newborn babies in developing countries has become the foremost priority to reduce the burden of RV-A gastroenteritis (RVGE) and to prevent childhood mortality. Thus, continuous efforts have been made to improve vaccine safety and efficiency. Two rotavirus vaccines, RotaTeq®, a three-dose pentavalent human-bovine reassortant strain (WC3, Merck and Co, NJ, USA) and Rotarix®, a two-dose monovalent live-attenuated human strain 89-12(P[8]G1) (GlaxoSmithKline, NC, USA) are licensed worldwide (Ruiz-Palacios et al. 2006, Vesikari et al. 2006) and Brazil initiated universal immunization in March 2006 with the oral vaccine, Rotarix®. Rotarix® has been demonstrated to be effective for preventing severe RVGE in diverse regions such as Asia, Europe and Latin America (100%, 98.5% and 84.7% effective, respectively) (Vesikari et al. 2007, Linhares et al. 2008, Phua et al. 2009). New data reported in Africa also indicate significant efficacy (61.2%) against severe RVGE (Madhi et al. 2010) and data from immunization surveillance in Brazil have demonstrated an important reduction in childhood hospitalization with severe RVGE (Leite et al. 2008, Carvalho-Costa et al. 2009, Gurgel et al. 2009, O'Ryan & Linhares 2009). Despite the fact that the G1P[8] vaccine is monovalent, it provides protection against a broad range of circulating strains, including strains carrying the G2 and the emerging G8 and G12 genotypes (O'Ryan et al. 2009, Sharma et al. 2010). Apparently, the noted significant shedding of this vaccine strain may indicate a high likelihood of transmission from vaccinated individuals to unvaccinated contacts (Salinas et al. 2005). Immune evasion by RV-A strains with G and/or P-types that are different from those present in current vaccines is a subject of discussion and concern. With this information, studying the comparative genomics of vaccine strains and immune-evading strains may help to identify genes encoding putative virulence factors that should be targeted to better understand vaccine escape mechanisms or for successful new vaccine development. Therefore, the aim of this study was to develop a protocol capable of rapidly differentiating the Rotarix® vaccine strain from wild-type G1P[8] strains in RV-A-positive samples. We describe a new approach based on in silico observations and restriction endonuclease digestion. Viral dsRNA from four faecal samples, genotyped as G1P[8] according to the Regional Reference Center for Rotaviruses of the Laboratory of Comparative and Environmental Virology, and from three different Rotarix® vaccine lots (#AR0LA029CA, #AR0LA040AA, #XR0LA055B1) were extracted using the glass powder method (Boom et al. 1990). The genes that code for the structural proteins (VPs: VP1, VP2, VP3, VP4, VP6 and VP7) and non-structural proteins (NSPs: NSP1, NSP3, NSP4 and NSP5) were amplified by reverse transcription-polymerase chain reaction (RT-PCR) according to previously described methods (Both et al. 1984, Das et al. 1994, Nakagomi & Kaga 1995, Matthijnssens et al. 2006a, Varghese et al. 2006, Tort et al. 2010). The PCR amplicons were purified and sequenced using an ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit® and an ABI Prism 3730 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA), as described by Otto et al. (2008). The chromatograms were analyzed using BioEdit® (Hall 1999). The NSP3 nucleotide sequences from clinical stool samples were entered into the GenBank/EMBL/DDBJ database under the following accessions: sp30843-86 (FN870369), rj7363-03 (FN870370), se16537-09 (FN870371) and se16897-09 (FN870372). The Rotarix® NSP3 gene sequence and accession will be released as soon as the National Program of Immunization authorises. The NSP3 vaccine nucleotide sequences from all segments (VPs and NSPs) were compared to the sequences from the reference strains and to other RV-A sequences available in GenBank using BLAST software (data not shown). There are two primary conditions that can be used to differentiate two viral strains, e.g. the vaccine strain from wild-type G1P[8] strains, by restriction enzymes: (i) a specific gene fragment that has mutations and allows for differentiation between the two strains and (ii) the stability of mutations over time. Thus, based on sequence alignment observations and the use of the NEBcutter program server (New England Biolabs) to test different restriction endonucleases, the NSP3 gene was selected from among the RV-A genes analyzed to be used as the marker due to the presence of a unique conserved point mutation resulting in the presence of a unique restriction site. All available data for the NSP3 gene in GenBank were retrieved and a total of 239 sequences were selected on April 7 2010. In silico digestion with a BspHI restriction enzyme resulted in two fragments (464 bp and 598 bp) specific for the Rotarix® vaccine strain. Fig. 1 shows the alignment of the vaccine NSP3 sequence and same NSP3 sequences from representative strains, demonstrating this particular difference. Although a large variety of restriction endonucleases can digest DNA at more than 150 recognition sites (Roberts et al. 2003) and other enzymes are capable of differentiating between rotavirus strains, only BspHI can achieve this differentiation in one digestion round. The length of the restriction fragments estimated in silico was confirmed using four clinical samples. The NSP3 amplicons from these samples were digested with the BspHI restriction enzyme and were compared with the BspHI-digested NSP3 vaccine amplicon, as shown in Fig. 2. None of the RT-PCR amplicons from the wild-type G1P[8] clinical samples were cleaved by BspHI. The 464-bp and 586-bp fragments were present in the digested RT-PCR amplicon from the vaccine strain, indicating a good correlation between the in silico and experimental data. The detection and identification of specific genotypes of RV-A in human stool samples can be performed using various methods, including type-specific PCR (Matthijnssens et al. 2006a), PCR amplicon length polymorphism (Gentsch et al. 1992), sequence analysis (Iturriza Gómara et al. 2002), capture and primer expression (Baggi & Peduzzi 2000, Barman et al. 2004, DiStefano et al. 2005) and hybridization to oligonucleotide probes (Lovmar et al. 2006). Our novel assay, based on the endonuclease restriction enzyme method, is a rapid and sensitive technique to distinguish the Rotarix® vaccine NSP3 gene from wild-type NSP3 genes in RV-A G1P[8] strains. The greatest advantages of this protocol are the lower risk of contamination and the lower costs compared to other published methods. Additional advantages are the reductions in time and manpower required when compared to RT-PCR followed by reverse hybridization (Leite et al. 1996, Chizhikov et al. 2002, Santos & Hoshino 2005, Santos et al. 2008, van Doorn et al. 2009). Similarly to the reverse hybridization method, our method discriminates between wild-type and vaccine strains based on a single-nucleotide change. Despite the fact that position 605 of the VP7 gene is a sequence signature for the vaccine strain (van Doorn et al. 2009), no marketed restriction enzyme is specific for this site. The BspHI enzyme is a low-cost restriction enzyme that hydrolyses position 464 of the NSP3 gene, allowing this digestion to be used a marker for the differentiation of wild-type G1P[8] strains from the vaccine strain. Nevertheless, the limitation of this method is that it does not provide information about the G and P types; thus, conventional RT-PCR is still needed to identify the genotypes. Although genetic drift is probably a slow process in rotavirus evolution, genetic shift through gene reassortment may be rather common and may explain the existence of similar genes in strains derived from animals and humans (Desselberger 1996, Parra et al. 2004, Matthijnssens et al. 2006b). These reassortment events could have a strong impact on RV-A ecology because of the potential to escape the immune response after vaccination. As described in other studies, the new approach shown here analyses a single gene segment; thus, tracking of the vaccine strain can be compromised. We suggest using a set of methodologies for exact vaccine discrimination and the proposed method will improve vaccine segregation accuracy because it is not based on the generally used VP4 and VP7 genes. Vaccine virus shedding and transmission to unvaccinated individuals could stimulate an immune response and confer protective immunity. As a result, the potential risk-to-benefit ratios of vaccine shedding among different groups and individuals are of special interest in low-income areas where reduced immunization rates exist but where high morbidity and mortality from RV-A occur (Anderson 2008). It is known that shedding is not associated with an increase in gastroenteritis-like symptoms and the rates of adverse reactions to the Rotarix® vaccine observed in clinical trials cannot be directly compared to the rates observed in clinical trials of any other RV-A vaccine. Additionally, adverse reaction rates may not reflect the rates observed in clinical practice. As with any vaccine, there is the possibility that the broad use of Rotarix® will reveal adverse reactions not observed in clinical trials (O'Ryan & Linhares 2009), thus emphasizing the importance of global surveillance. Monitoring the genotypes and frequency of circulating strains in a community will allow the determination of how routine immunization impacts the prevalence of common genotypes, the emergence of immunity-escaping strains and the evolution of wild-type rotavirus genes as a consequence of immune selective pressure. Because evolutionary changes may occur constantly in different gene segments, resulting in genetic drift of the vaccine strain from the prototype strain, supervising vaccine evaluation is also a critical part of surveillance programs. Variations must always be ascertained because genetic fluctuations may theoretically lead to reduced vaccine efficacy or even to vaccine failure over time. Therefore, the continuous surveillance of circulating strains is required and research on the evolutionary rate of different gene segments is important for evaluating and updating vaccine analysis methods. ACKNOWLEDGEMENTS To Rosane Maria Santos de Assis and Alexandre Madri Fialho, for technical assistance, to Edson Pereira Filho, from The Army Biology Institute (IBEx), and Marli Costa, from CGlab, to provide vaccine samples, to Maria Eugenia Galeano, for helpful comments, to mothers who agreed that their children participate in this study, to the private and state hospitals, for providing samples for the development of this study, and to the PDTIS DNA Sequence Platform staff at Fiocruz (RPT01A), for his technical support in sequencing reactions. REFERENCES
Copyright © 2010 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc10184f2.jpg] [oc10184f1.jpg] |
| |||||||||

{kind=link}
{kind=link}