 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 1, 2011, pp. 16-22 ARTICLES Feeding behavior of Triatoma vitticeps (Reduviidae: Triatominae) in the state of Minas Gerais, Brazil Rita de Cássia Moreira de SouzaI; Adriana C SoaresII; Ceres Luciana AlvesII; Elias Seixas LorosaIII; Marcos Horácio PereiraII; Liléia DiotaiutiI, + ILaboratório
de Triatomíneos e Epidemiologia da Doença de Chagas, Instituto
René Rachou-Fiocruz, Av. Augusto de Lima 1715, 30190-002 Belo Horizonte,
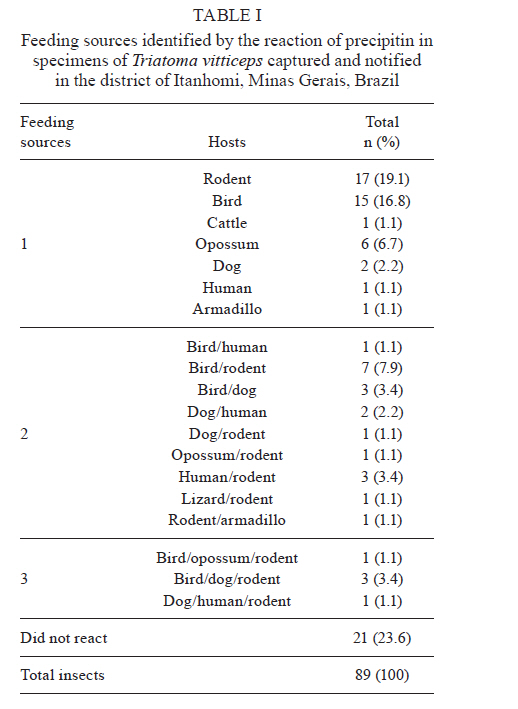
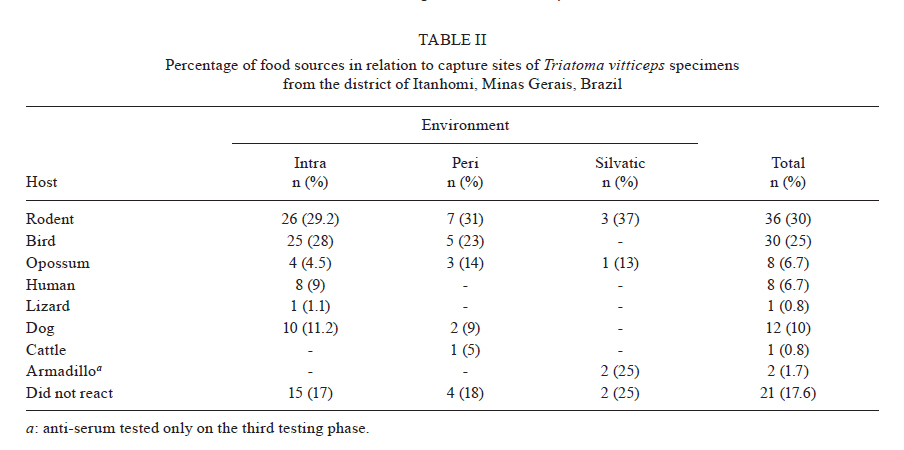
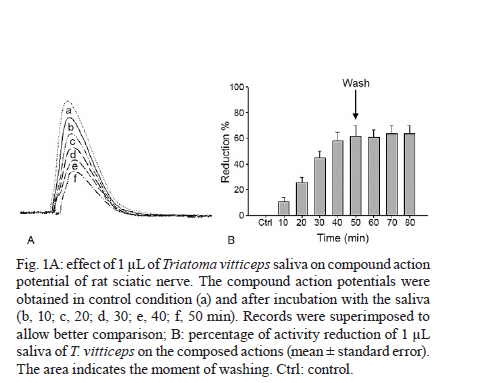
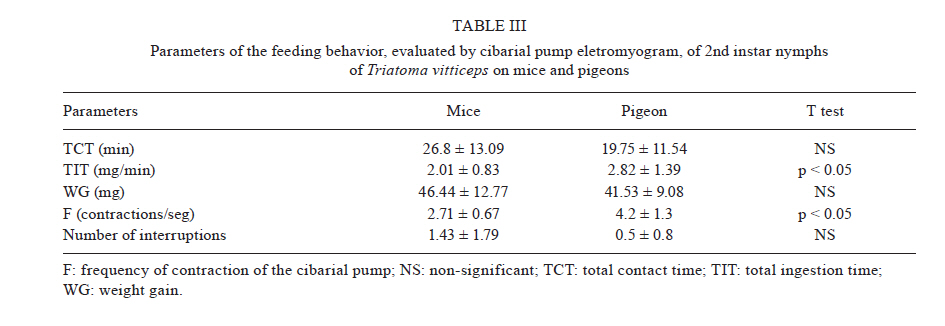
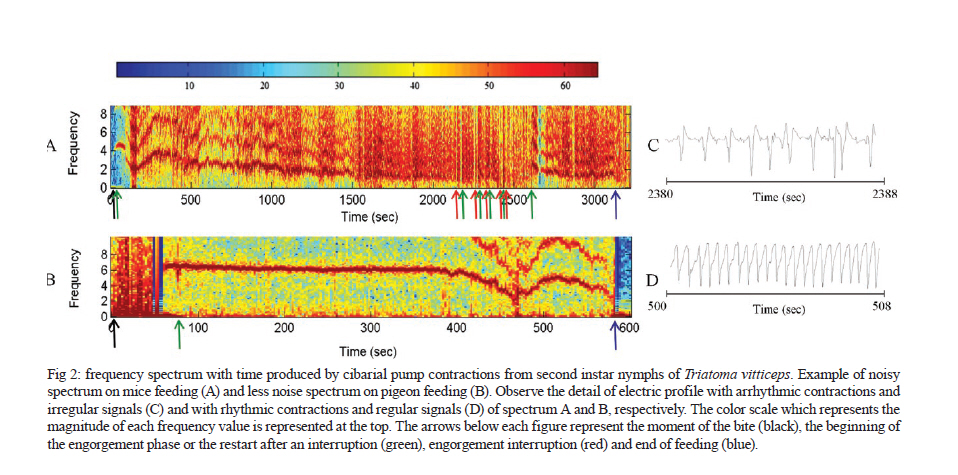
MG, Brasil Received 5 May 2010 Financial support: UNDP/World Bank/WHO/TDR, FIOCRUZ, CNPq, FAPEMIG Code Number oc11003 ABSTRACT The objective of this study was to evaluate the feeding behavior of Triatoma vitticeps through the identification of its food sources and the characterization of the blood ingestion process. In addition, we aimed to verify if the saliva of this vector interferes with the perception of the host during the feedings by creating a nervous impulse. Here, we demonstrated that the T. vitticeps saliva reduces, gradually and irreversibly, the amplitude of the compound action potential of the nervous fibre, which helps decrease the perception of the insect by the host. The precipitin reaction demonstrated the feeding eclecticism of this vector, with the identification of eight food sources - most of them found simultaneously in the same insect. The analysis of the electrical signals produced by the cibarial pump during meals demonstrated that the best feeding performance of T. vitticeps nymphs that fed on pigeons is mainly due to the higher contraction frequency of the pump. The longer contact period with the host to obtain a complete meal compared with other triatominae species of the same instar could favor the occurrence of multiple blood sources in T. vitticeps under natural conditions, as it was evidenced by the precipitin test. Key words: hemiptera - feeding behavior - saliva - cibarial pump - hematophagic Triatoma vitticeps is a triatominae species found in the Brazilian states of Espírito Santo (ES), Minas Gerais (MG), Rio de Janeiro (RJ) and the southernmost regions of Bahia (Lent & Wygodzinsky 1979, Silveira et al. 1984). Studies carried out on T. vitticeps populations in ES, RJ and MG demonstrated the capacity of this triatomine to invade houses (Santos et al. 1969c, Ferreira et al. 1986, Sessa & Carias 1986, Dias et al. 1989, Gonçalves et al. 1998, Souza et al. 2008). Similarly, these invasive insects were always highly infected with flagellates similar to Trypanosoma cruzi (Pinto et al. 1969, 1986, Santos et al. 1969a, b, Barros et al. 1975, Sessa & Carias 1986, Dias et al. 1989, Gonçalves et al. 1998, dos Santos et al. 2005, 2006). On the other hand, colonization is not efficient and several studies on the feeding habits of this triatomine using the precipitin method demonstrated some difficulty to obtain a full meal. The life cycle and population dynamics of triatominae species are directly related to their interaction with vertebrate hosts (Schofield 1994). The host's ability to perceive the insect during the feeding process represents a threat to the insect and may determine its success in obtaining blood. When the vertebrate host notices the presence of the insect, it attempts to eliminate it mechanically, which can cause an early interruption of the feeding or even the insect's death (Rossignol et al. 1985). Species in the subfamily Triatominae minimize such a threat with two cooperative mechanisms: the production of a salivary anaesthetic (Dan et al. 1999) and the development of an efficient muscular pump (the cibarial pump), which is capable of generating high pressures that allow the ingestion of large quantities of blood in a short period of time (Bennet-Clark 1963). In these vectors, the contact time with the hosts is influenced by the blood ingestion rate, which varies among triatomine species, the developmental stage and the physiology of the host (Guarneri et al. 2000, 2003, Sant'Anna et al. 2001). The identification of the food source of hematophagous insects is highly used for epidemiological studies of infectious diseases to find the vectorial potential presented by the insects in various diseases, such as the Chagas disease. In 1960, Siqueira (1960) demonstrated that it was possible to find, through the precipitin reaction, the feeding source of triatomines kept under laboratory settings, even when the blood ingestion had occurred 120 days earlier. Using this technique, Dias et al. (1989), Gonçalves et al. (2000) and Lorosa et al. (2003) demonstrated that T. vitticeps have a significant feeding eclecticism, a fact of great epidemiological importance because a T. cruzi infection may derive from different sources. The occurrence of multiple blood meals in T. vitticeps might be the result of the unfeasibility to obtain a full meal from a single source. Therefore, information about how the perception of the host can interfere with the feeding process of T. vitticeps would be fundamental to understand the aspects related to the feeding behavior of T. vitticeps. In addition, the identification of the feeding sources, the dynamics of blood ingestion and the generation of in vitro nerve impulses triggered by the vector's saliva to assess the interference of the hosts' perception are of crucial importance. MATERIALS AND METHODS The insects - The T. vitticeps used in this study derived from the Itanhomi district (19º11'15"S, 41º48'45"W), located at 387 km from Belo Horizonte, MG. For the identification of the feeding sources, insects captured by health agents were used, as well as insects notified by the local population. For the study of the saliva and the monitoring of the cibarial pump, the insects were reared in the laboratory under controlled conditions of temperature (27 ± 2ºC) and humidity (60 ± 10%). Identification of the feeding sources - The 89 specimens of T. vitticeps analyzed were adults. Of these, 82 were derived from the intra and peridomicile areas (barns and basements), while seven were captured from light traps in the silvatic environment through luminous trap. Samples of the stomach contents of live insects were directly impregnated in filter paper. When the insects were dead and the stomach contents were dried, they were dissected and the extracted material was eluted in 1 mL of saline solution before its absorption with filter paper. After the papers were dried, they were wrapped in aluminium foil and stored individually at -20ºC. For the precipitin test, the antisera preparation, the titers determination and the specificity were performed according to Lorosa et al. (1998). The blood sources investigated were the probable hosts that occur in the study area. We used the following antisera: human (1:15.000), rodent (1:15.000), bird (1:10.000), opossum (1:12.000), lizard (1:8.000), dog (1:15.000), cattle (1:13.000) and armadillo (1:10.000). Action of T. vitticeps saliva - The saliva was individually collected with a capillary tube by the mechanical stimulation of adult insects and 5th instar nymphs 15 days after feeding. We used 30 insects to extract saliva. The saliva discharged was transferred to a microtube and kept in ice to form a pool of saliva. The extraction was performed on the day before the experiment and it was maintained at -20ºC until use. The experiments were performed on the sciatic nerve of the Wistar strain and the compound action potentials were recorded using the single sucrose-gap method, as described by Cruz et al. (1994). The nerves were carefully removed, desheathed and placed across the five compartments of the experimental chamber, on a narrow groove sealed with vaseline to isolate the compartments and to mechanically secure the nerve bundle. The first two compartments were used for supramaximal stimulation at a frequency of 0.1 Hz. The central compartment, containing 200 μL of the test solution, was grounded and electrically isolated from the 5th (recording) compartment through which a 282 mM sucrose solution constantly flowed. The other compartments were filled with Locke's solution, which contained (in mM): 154 NaCl, 5.5 kCl, 2.2 CaCl2, 5.0 N- (2-hydroxyethyl) piperazine-N'- [2-ethanesulphonic acid (HEPES)], 5.0 glucose and the pH was adjusted to 7.5 using NaOH. Compound action potential amplitudes > 35 mV were routinely recorded and remained stable for more than 5 h after stabilization of the recording conditions (± 1 h). The electrodes placed in the test and the 5th compartments were fed into a differential amplifier with fixed gain (50 times). The resulting electrical signal was then collected via a digital data acquisition plate (ADC100®-Pico Technology Limited, UK) and stored in an IBMAT compatible microcomputer for subsequent off-line analysis. Feeding behavior - For this experiment, we selected 2nd instar nymphs of T. vitticeps with approximately 12 days of fasting after molting. This instar was selected because of its shorter time to obtain a full meal. Pigeons and hairless mice were used as feeding sources. The mice were anesthetized intraperitoneally with a mixture of Ketamine and Xylazine (Agener União; 150 and 10 mg/kg, respectively). Twenty insects were individually allowed to feed on the abdominal region of the mice. For the pigeon assays, the birds were immobilized and 20 insects were individually allowed to feed on a small area of the breast from which all feathers had been removed. Care was taken to minimize light, odor, noise and vibrations when handling the insects before testing. The feeding process was analyzed through a monitoring system of the electrical activity of the insects' cibarial pump developed from the Smith's technique (Smith 1979) and modified by Guarneri et al. (2003). One electrode was fixed to the host and a 2nd electrode was connected to a metal mesh, which was fixed to a container used as substrate for the insects. These two electrodes detected the electrical signals from the cibarial pump. The signals were filtered to reject frequencies above 17 Hz (with a slope of 45 dB/octave), amplified 200 times and collected via a digital data acquisition plate (ADC100®-Pico Technology Limited, UK) connected to an IBMAT compatible microcomputer. The electrode and the metal mesh were moistened with an electrolytic gel (Regisgraf-Gel®) to facilitate the passage of the electrical signals. To determine weight gain, the insects were weighed immediately before (initial weight) and after feeding. Each insect was allowed to feed until it removed its mouthparts without trying to probe again within 1 min. The engorgement phase was considered to start after at least 10 consecutive characteristic peak-like signals attributed to the cibarial pump contractions. From the cibarial pump records, the total contact time was defined as the time during which the insect mouthparts remained inserted into the host skin. The non-ingestive time, the initial cumulative probing time and the interruption time were also determined. The non-ingestive time was defined as the time when insects were not pumping, so probing time plus any interruption to feeding comprised this time. The cumulative probing time was defined as the sum of the times from the 1st insertion of the mouthparts into the host up to the beginning of the engorgement phase. If the bug ended an initial probing and restarted another one elsewhere, the 1st probing time was added to the 2nd and so successively. The last probing time was the time from the last bite until the start of the engorgement phase. The interruptions were defined as stops for more than 4 s in blood pumping after the engorgement phase had begun. The interruption time was defined as the sum of all the individual interruption times. The pump frequency was defined as the total number of cibarial pump contractions divided by the effective contact time (i.e., the time in which the pump was effectively working, without probing or interruptions). The total ingestion rate was calculated by dividing the weight gain by the total contact time and the effective ingestion rate by dividing the weight gain by the effective contact time. The number of bites per insect was also counted. Software that works under a Matlab (version 6.5.0) environment was developed to analyze the records of the electromyogram of the cibarial pump. This software enables the analysis of the frequency spectrum from the signals generated by the cibarial pump in relation to the time of feeding (Araujo et al. 2009). The results were analyzed with the Kolmogorov-Smirnov test to evaluate if the distribution of each variable was normal. The variables were then compared through the Student t test. The statistical significance was determined at p < 0.05. RESULTS Study of the feeding source - The results of the blood meal identification were based on 89 specimens. From these, 21 were no reactive to the antisera tested, 43 (48.3%) reacted to a single source, 20 (22.5%) reacted to two sources and five (5.6%) reacted to three different sources. When the insects fed on more than one host, two or more sources of blood could be found in a single insect sample. Our data suggest that the most frequent association, considering all the samples, was birds and rodents blood (7.9%) (Table I). By grouping the insects that fed on a single source with those that fed on two or more, the rodents' blood prevailed, totalizing 36 samples (30%), followed by birds' blood (25%), found in 30 of the processed samples (Table II). The origin of the ingested blood and the percentage of positive reactions to its respective antisera are shown in Table II. The identification of the blood meals showed that the rodents are the main food source in the three studied environments. The birds were the 2nd most common food source in domestic and peridomestic habitats. In the forested surroundings, the armadillo was also a feeding source for this vector. However, its anti-serum was only available for part of the insect samples. Human blood appears only at the intra-domicile area with 9% of the reactions. Action of the saliva of T. vitticeps on the generation of nerve impulses - The addition of 1 μL of T. vitticeps saliva in the preparation of the nerve produced a progressive reduction of the amplitude of the compound action potentials, without showing alterations on the repolarization phase when compared to the control. Fig. 1A illustrates one of the four replicas of this experiment. No recovery of the compound action potential amplitude was observed even 30 min after washing the preparation with Lock's solution. The mean reduction observed was 66.1% ± 6.3 (n = 4) (Fig. 1B). Dynamics of blood ingestion: electrical monitoring of the cibarial pump - The nymphs of T. vitticeps that fed on mice showed a lower mean blood ingestion rate than the rate observed in pigeons and it was mainly due to the lower contraction frequency of the cibarial pump. The weight gain of the insects was similar with both hosts, whereas the total contact time and the number of interruptions tended to be higher when the insect fed on mice (Table III). Feeding on mice was more difficult for the insects as evidenced by the rhythm of contractions and shape of the electrical signals generated by the cibarial pump during the feeding process. When the contractions were arrhythmic and the electrical signal showed irregular shape (2 peaks or intermediary forms between 1 or 2 peaks) a strong noise is observed in the graph of the frequency spectrum through time. This type of signal was constant in 72.7% of the experiments done on mice (Fig. 2A). On the other hand, 81.8% of the cibarial pump measurements from nymphs feeding on pigeons contracted rhythmically and generated regular electrical signals (single peak signals) that generated lower noise on the frequency spectrum through time, as shown in Fig. 2B. DISCUSSION The feeding process of triatomine bugs greatly varies among species and it is influenced by various factors, such as the characteristics of the feeding apparatus and the composition of the saliva, as well as the physiology of the host and the feeding site (Guarneri et al. 2000, 2003, Araujo et al. 2009). Therefore, the capacity to explore available food resources can directly influence the population dynamics and the vectorial capacity of triatominae (Pereira et al. 2006). Having saliva that reduces the noxious sensation is advantageous because the quantity of blood obtained by triatomines is limited by the irritation caused during the feeding process (Pereira et al. 1995, 1998). Dan et al. (1999) observed that the saliva of Triatoma infestans has an irreversible inhibitory activity on the generation of nervous impulses and this effect may decrease the sensitivity of the region where the insect probes. It has also been demonstrated that Panstrongylus megistus has an inhibitory activity on the generation of nervous impulses (Alves 2007). However, these two triatominae species differ in the reversibility of the inhibition because, contrary to what was observed for T. infestans, the saliva of the P. megistus produced a progressive inhibition in the amplitude of action potential in a reversible manner. Our results are very similar to those described for the T. infestans saliva. No recovery of the compound action potential amplitude was observed even 30 min after washing the preparation with Locke's solution. This lasting effect might be explained by the high affinity of the active principle of the saliva to the sodium channel or by a similar effect to that observed in response to high concentrations of local anesthetics, which cause a nonreversible block in isolated nerves (Bainton & Strichartz 1994, Lambert et al. 1994). Thus, we demonstrated that the T. vitticeps saliva acts on the nerve endings by inhibiting the generation of nerve impulses and, consequently, the perception of the insect by the host. Studies on the identification of food sources leads to the discovery of possible natural reservoirs and evaluates the degree of movement of the parasite in the area, contributing to a better understanding of the local epidemiology. Similarly, these studies are important because the natural ecotopes of T. vitticeps are unknown. T. vitticeps captured in different sites of ES and RJ showed that the feeding habits of this triatominae species are eclectic (Dias et al. 1989, Gonçalves et al. 2000, Lorosa et al. 2003). The results presented by Dias et al. (1989) showed a marked anthropophilia, with 89.2% of the antisera reacting to human types, even in the absence of colonization. Our results are pioneers in identifying the food source of T. vitticeps from MG. Again, these insects showed a wide eclecticism in their feeding habits with high incidence of rodents (30%) and birds (25%). During the insects capturing, the presence of rodents inside homes was indirectly verified by the observation of feces and by reports of the inhabitants. Besides that, the district was once endemic for the bubonic plague and these synanthropic mammals are abundant in wild environments, which are close to the urbanized areas. Chickens are the most frequent domestic animals and move freely around the houses' backyards, which could have contributed to the results found by the precipitin method. Moreover, considering that the infection is acquired outside of the home environment, the birds are refractory to infection and that there is an intense exchange between wild and artificial environments (Souza et al. 2008), these results suggest a close relationship between T. vitticeps and rodents. Other domestic animals are potential feeding sources of T. vitticeps, such as dogs (10%). These animals are mentioned as important hosts for T. cruzi in various countries of the American continent (Barretto et al. 1980), with Argentina being characterized as the main domestic reservoir (Wisnivesky-Colli et al. 1985, Cohen & Gürtler 2001). Therefore, dogs have an important epidemiological role for the Chagas disease because the infection of these animals can be a potential link between the domestic and the wild cycles of the disease. Despite the absence of intradomiciliary colonies, the presence of human blood in the digestive tube of the T. vitticeps specimens captured in this environment showed that triatominae adults that invade houses are able to feed on their inhabitants. This highlights the maintenance of an efficient system of entomologic surveillance. In addition, the multiple blood meals found reinforce the fact that this species circulates between wild and artificial environments showing high rates of infection by T. cruzi. The analysis of the electrical activity of the cibarial pump during the feeding process of T. vitticeps nymphs allowed the monitoring of its functioning. Similarly, we identified several events such as the bite and the beginning of the exploring phase, the contraction of the pump's dilating muscle, the interruptions of blood pumping and the removal of mouthparts from the host's skin at the end of the feeding. This information and the data on weight gain of the insect allowed the calculation of several parameters of the feeding process. The duration of the contact is an important factor in the interaction between hematophagous arthropods and their hosts because the defensive behavior of hosts can reduce the feeding success or even cause the death of the insect (Guarneri et al. 2003). Therefore, species that possess higher total ingestion rates tend to achieve higher nutritional status and reach higher population densities as occurs in T. infestans and Rhodnius prolixus, the most important vectors of T. cruzi in South and Central America, respectively (Sant'Anna et al. 2001). Our data demonstrated that the nymphs of T. vitticeps demand a longer contact time when they feed on mice. According to Guarneri et al. (2000) and Sant'Anna et al. (2001), when nymphs of the same species and the same developmental stage feed on different hosts, the variations in contact time are mainly related to differences in the pump frequency and the number of interruptions. In the present study, the lower blood ingestion rate in mice is mainly related to the lower frequency of contractions of the cibarial pump in this host compared to the pigeon (2.7 and 4.2 contractions/s, respectively). However, the results presented by the precipitin reaction demonstrate that, in natural conditions, both birds and in rodents are important food sources for T. vitticeps and that the feeding difficulties are not an obstacle to the use of rodents. The blood ingestion rate of T. vitticeps 2nd instar nymphs (2.0 mg/min) was higher than that reported for the same developmental stage of Triatoma brasiliensis and Triatoma pseudomaculata (1.0 and 0.4 mg/min, respectively) feeding on mice (Soares et al. 2000). However, T. vitticeps is a large triatominae that needs higher quantities of blood to complete its cycle; therefore, it considerably increases the contact time with its hosts. Comparatively, the contact time of T. vitticeps 2nd instar nymphs was higher (26.8 min) than that reported for T. brasiliensis and T. pseudomaculata (22.1 and 19.5 min, respectively) (Soares et al. 2000). During the feedings process of nymphs on mice and pigeons, it was possible to notice the difficulty in obtaining blood due to the irregularity of the electrical signals produced by the cibarial pump, both in frequency and in profile, generating noisy frequency spectra as a function of time. This was especially true in experiments with mice, if we compare the frequency spectra obtained during the R. prolixus nymphs feeding on mice when similar experimental conditions were followed (Araujo et al. 2009). Our results corroborate the current knowledge on the biology and the behavior of T. vitticeps. Despite this, it was demonstrated that the saliva of this vector decrease the local sensibility of the hosts. The difficulty in blood obtainment, mainly due to the low contraction frequency of the cibarial pump in mice, could partly explain the feeding eclecticism observed for this triatominae species. REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11003t1.jpg] [oc11003f1.jpg] [oc11003t2.jpg] [oc11003t3.jpg] [oc11003f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}