 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 2, 2011, pp. 139-145 ARTICLES Detection of rifampin-resistant genotypes in Mycobacterium tuberculosis by reverse hybridization assay Raquel de A MaschmannI, II, +; Mirela VerzaI, II; Marcia SN SilvaI, III; Rosa Dea SperhackeIV; Marta O RibeiroI; Philip Noel SuffysV; Harrison Magdinier GomesV; Enrico TortoliVI; Fiorella MarcelliVI; Arnaldo ZahaII; Maria Lucia R RossettiI, II, III IFundação
Estadual de Produção e Pesquisa em Saúde, Porto Alegre,
RS, Brasil + Corresponding author: raquel368@gmail.com Received 27 July
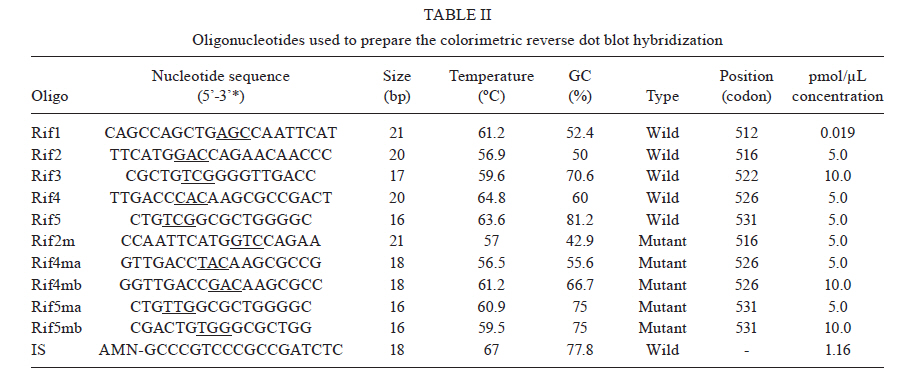
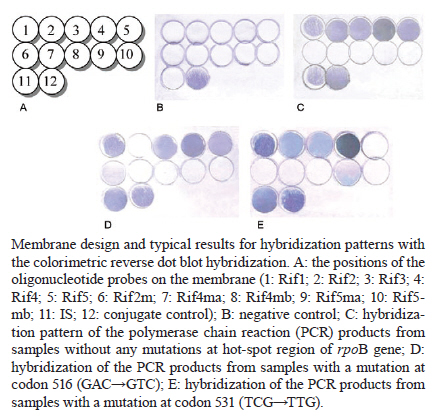
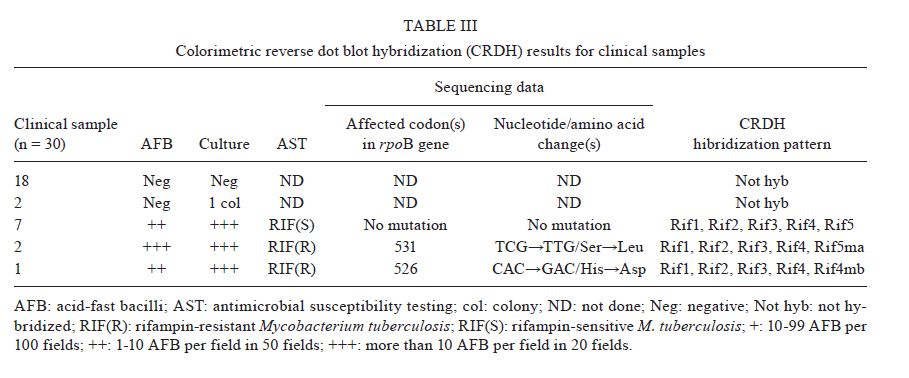
2010 Code Number: oc11023 ABSTRACT We used a colorimetric reverse dot blot hybridization (CRDH) assay to detect the presence of mutations in a specific region of the rpoB gene, associated with rifampin (RIF) resistance, in a panel of 156 DNAs extracted from 103 RIF-sensitive and 53 RIF-resistant cultures of Mycobacterium tuberculosis. When compared with the antimicrobial susceptibility test (AST), the sensitivity and specificity of the CRDH were 92.3% and 98.1%, respectively. When compared with sequencing, the sensitivity and specificity of the CRDH were 90.6% and 100%, respectively. To evaluate the performance of the assay directly in clinical specimens, 30 samples from tuberculosis patients were used. For these samples, the results of the CRDH were 100% consistent with the results of the AST and sequencing. These results indicate that the rate of concordance of the CRDH is high when compared to conventional methods and sequencing data. The CRDH can be successfully applied when a rapid test is required for the identification of RIF resistance in M. tuberculosis. Key words: drug resistance - reverse dot blot hybridization - rifampin - rpoB - tuberculosis Tuberculosis (TB) is a global health problem and approximately 1.5 million people die from TB every year. Brazil is one of the 22 countries responsible for 80% of the total global TB incidence. The estimated TB incidence in Brazil is 48 out of 100,000 inhabitants (WHO 2009). A new and potentially devastating threat to TB control is the emergence of strains that cannot be cured by standard anti-TB drug regimens (Lawn & Wilkinson 2006). Drug resistant TB commonly arises through the selection of mutated strains by inadequate chemotherapy. Resistance to at least two major anti-TB drugs, rifampin (RIF) and isoniazid (INH) has been termed multidrug-resistant TB (MDR-TB). Treatment of MDR-TB requires prolonged and expensive chemotherapy using second-line drugs. These problems compromised the progress made in TB control during the past decade (WHO 2009). Resistance to RIF is an excellent marker for MDR-TB, as 90% of RIF-resistant Mycobacterium tuberculosis strains are also INH resistant (Telenti et al. 1993, Cole & Telenti 1995, Ramaswamy & Musser 1998). Molecular tests for the detection of RIF resistance has also been developed because almost all RIF resistant M. tuberculosis strain mutations are found in the short hot-spot region (81-bp) of the rpoB gene (Kapur et al. 1994, Telenti et al. 1997, Valim et al. 2000), which encodes the β-subunit of the RNA polymerase (Telenti et al. 1993, Heep et al. 2001). In isolates from Brazil (Valim et al. 2000) and other countries (Hillemann et al. 2005, Miotto et al. 2006), the most frequently affected codons by point mutations are 531, 526 and 516. An accurate and early detection of drug resistance in clinical M. tuberculosis isolates is crucial for the appropriate treatment and to prevent the development of further resistance and the spread of resistant strains. Drug susceptibility testing by conventional methods takes more than four weeks. Generally, DNA sequencing-based approaches are considered the reference assays for the detection of mutations, but often they have been found to be too cumbersome for routine use. The commercial molecular hybridization tests, such as GenoType MTBDR (Hain Lifescience, Nehren, Germany) and INNO-LiPA Rif.TB (Innogenetics, Zwijndrcht, Belgium) are sensitive and specific, but the high cost and the need for validation hamper the widespread application of the test-kits in geographic areas where it is most urgently needed (Mäkinen et al. 2006). In this study, we tested a molecular assay for the detection of M. tuberculosis RIF resistance based on colorimetric reverse dot blot hybridization (CRDH). Previously, a similar strategy was developed by Verza et al. (2009) in our laboratory. The results from the CRDH were compared with the data obtained by DNA sequencing and antimicrobial susceptibility test (AST) and some DNAs were also analyzed by the GenoType MTBDR commercial test. MATERIALS AND METHODS Strains and clinical isolates - To standardize and evaluate the CRDH, we used a panel of 156 DNAs obtained from M. tuberculosis cultures that had been submitted to both AST and partial rpoB gene sequencing. This panel contained 103 DNA samples with no mutations and 53 DNA samples with one or more mutations in the hot-spot region of the rpoB and associated with RIF-resistance in this region (Table I). The strains were selected from the collection of the Institute of Biological Research/Central Laboratory of Rio Grande do Sul, Brazil (IPB/LACEN-RS). The M. tuberculosis H37Rv (ATCC 27294) reference strain was used as a wild-type (WT) control. The specimens had previously been cultured on Ogawa medium and identified as M. tuberculosis based on a positive niacin test. AST was performed using Lowenstein-Jensen medium according to the proportion method (Canetti et al. 1963). After standardization with DNA extracted from the cultures, the CRDH method was performed on DNA extracted from 30 clinical isolates from the IPB/LACEN-RS, Brazil, and the Laboratorio di Microbiologia e Virologia of the Centro Regionale di Riferimento per i Micobatteri, Ospedale di Careggi, Florence, Italy. These samples were examined microscopically for the presence of acid-fast bacilli and then submitted to AST and partial rpoB gene sequencing. DNA extraction - Nucleic acids were extracted from M. tuberculosis cultures using the CTAB method as described by van Soolingen et al. (1994). Nucleic acids of M. tuberculosis from the clinical samples were extracted using the method adapted from Boom et al. (1990). All respiratory specimens had been decontaminated previously by the N-acetyl-L-cysteine-NaOH conventional method (Kent & Kubica 1985). Polymerase chain reaction (PCR) amplification - For development of the CRDH, a 157-bp fragment of the rpoB gene was amplified using the primers RF1 (5'-GGTCGCCGCGATCAAGGAGT-3') and RF2 (5'-TGCACGTCGCGGACCTCCA-3') and a 245-bp fragment of the insertion element sequence IS6110 (TB control) using the primers INS1 and INS2 as described by Hermans et al. (1990). The reverse primers were biotin-labelled and the PCR-duplex was standardized in a final volume of 50 μL containing 200 μM of each dNTP, 10 mM Tris-HCL (pH 8), 50 mM KCL, 2 mM MgCl2, 10 pmoles of each primer (RF1/RF2 and INS1/INS2), 50-100 ng of purified DNA and 2.5 U of Taq DNA polymerase (Cebiot, UFRGS, Brazil). The PCR reactions were carried out as follows: 3 min at 95ºC, 30 cycles of 1 min at 95ºC, 1 min at 60ºC, 1.5 min at 72ºC, and 4 min at 72ºC. For amplification of the clinical samples, 10 μL of the processed material was submitted to the same conditions as above for 40 thermal cycles. The PCR products were analyzed by electrophoresis in a 1.5% agarose gel. DNA sequencing of the rpoB gene - For sequencing, a 157-bp fragment of the rpoB gene was amplified with the primers, RF1 and RF2, as described above. Amplifications were carried out as follows: 30 cycles at 94ºC for 2 min, 65ºC for 1 min and 72ºC for 2 min. The PCR products were purified using the polyethylene glycol method, listed on the Multilocus Sequence Typing website (http://pubmlst.org/neisseria/mlst-info/nmeningitidis/pcr.shtml) and sequenced using Big Dye® Terminator Cycle Sequencing Kit with AmpliTaq DNA polymerase (Applied Biosystems, Foster City, CA, USA) in the ABI Prism 3100 DNA Sequencer (Applied Biosystems). The sequences were analyzed using programs PREGAP and GAP4 from STADEN package 10.0. Nucleotide sequences with phred values > 20 were considered for analysis. GenoType MTBDR assay - The CRDH results of 15 out of 156 DNAs were compared with the results of GenoType MTBDR, a molecular test used for the determination of mutations associated with susceptibility to RIF and INH. The assay was performed according to the manufacturer's instructions. CRDH test - A total of 10 oligonucleotide probes were designed to screen the most frequent WTand mutant genotypes of the rpoB gene using the Primer Express Software v2.0 (Applied Biosystems). Additionally, one oligonucleotide probe specific to M. tuberculosis complex (MTBC), complementary to an internal sequence of the insertion element IS6110, was constructed. All oligonucleotide probes (Invitrogen) were designed with a 5'-terminal amino group. The oligonucleotide sequences are listed in Table II. The negatively-charged nylon membrane (Biodyne C, Pall Corporation) was activated in a solution of 16% (w/v) EDAC [1-(3-dimethylaminopropyl)-3-ethylcarbodiimide hydrochloride, Acros Organics] for 15 min at room temperature (RT) and washed in distilled water for 2 min at RT. Then 10 μL each of oligonucleotide probe, diluted to the appropriate concentration (Table II) in 0.5 M NaHCO3 (pH 8.4) and the conjugate control (CC) (0.01 μg/μL of streptavidin-alkaline phosphatase conjugate, Invitrogen) were spotted separately in 12 circles and incubated for 1 min at RT. Membranes were incubated in 0.1 M NaOH for 10 min, washed in distilled water for 2 min at RT, washed in 2x SSC/0.1% SDS for 10 min at 50ºC and finally incubated in 20 mM EDTA (pH 8) for 15 min at RT. The membranes were then transferred into a polyethylene tube and stored at 4ºC for posterior use. The membranes were washed in 2x SSC/0.1% SDS for 5 min at 50ºC and incubated in 2x SSC with 3% bovine serum albumin (BSA) (INLAB) at 50ºC for 15 min. Twenty microliters of each PCR reaction was added to 150 μL 2x SSC/0.1% SDS, denatured at 100ºC for 10 min and transferred to an ice bath. The denatured PCR product was then added into polyethylene tube containing a membrane in 1 mL of hybridization solution (2x SSC/0.1% SDS). The hybridization was achieved by incubating at 62ºC for 45 min. After hybridization, the membranes were washed in 2x SSC/0.5% SDS and in 0.2x SSC/0.5% SDS at 57ºC for 10 min each. The membranes were then treated with a Tris-buffered saline (TSB) (100 mM Tris-HCl, 150 mM NaCl, pH 7.5) containing 6% BSA for 30 min at 50ºC, followed by a second incubation in TSB for 5 min at RT. The membranes were incubated in TSB buffer, containing 0.33 μg/mL streptavidin-alkaline phosphatase conjugate, for 15 min at RT. The unbound conjugate was removed by washing in TSB buffer for 10 min, followed by alkaline phosphatase buffer (AP buffer) (100 mM Tris-HCl, 150 mM NaCl and 5 mM MgCl2.6H2O, pH 9.5) for 10 min at RT. The hybridized DNA was visualized by adding 40 μg/mL 5-bromo-4-chloro-3-indoyl phosphate and 82.5 μg/mL nitro blue tetrazolium (BCIP/NBT, Sigma) in AP buffer and incubated for 10 min at RT. A purple precipitate was obtained when there was a perfect match between the probe and the biotinylated PCR product. The colorimetric reaction was blocked with distilled water, the membranes were then dried at RT and the results were recorded. The whole procedure was also performed for the positive and negative controls. Determination of sensitivity and specificity of the CRDH - The analytical sensitivity of the CRDH was determined by the procedure described above with a serial 10-fold dilution of the M. tuberculosis H37Rv DNA (100 ng-1 pg). To determine the specificity of the CRDH, 100 ng of genomic DNA from each of the following strains was submitted to the entire procedure: Neisseria meningitidis, Streptococcus pneumoniae, Salmonella enterica, Haemophilus influenzae, Escherichia coli (obtained from IPB/LACEN-RS), Mycobacterium marinum, Mycobacterium intracellulare, Mycobacterium scrofulaceum, Mycobacterium gordonae, Mycobacterium avium, Mycobacterium smegmatis, Mycobacterium kansasii, Mycobacterium xenopi, Mycobacterium fortuitum-peregrinum and Mycobacterium phlei (obtained from Fiocruz, Brazil). RESULTS Comparision of CDRH with the proportion method - Five WT probes Rif1, Rif2, Rif3, Rif4 and Rif5 were used to detect the hot-spot region of the rpoB in the M. tuberculosis, including the sequence from codon 509-534. The isolates that showed positive signals only with WT probes were interpreted as being sensitive to RIF (Figure). The results for a mutant genotype demonstrated the lack of hybridization on one of the five WT probes and/or hybridization on one or more of the mutant probes (Rif2m, Rif4ma Rif4mb, Rif5ma or Rif5mb) as shown in Table II. According to the AST, 52 resistant strains were detected. Out of these strains, four strains were not detected correctly by the CRDH. The first strain had mutations at codons 511 (GTC'!CGC) and 516 (GAC'!TAC) with a weak hybridization signal at the Rif1 and Rif2 respectively. The second strain had a mutation at 516 (GAC'!TAC) with a weak signal at the Rif2. The third strain had a mutation at 513 (CAA'!CCA) with a weak signal at the Rif1. All three strains were confirmed by sequencing. Finally, the fourth isolate hybridized with all WT probes. The results from the AST showed that 102 (98.1%) out of 104 M. tuberculosis strains were RIF-sensitive and hybridized only with the WT probes. Of the remaining two, one had a mutation at codon 531 (TCG'!TGG) and another had a mutation at the Rif4 region. When compared with the conventional AST, the sensitivity and specificity of the CRDH were 92.3% and 98.1%, respectively. Comparision of CRDH with DNA sequencing - Out of 53 M. tuberculosis strains that had mutations in the hot-spot region of the rpoB gene according to sequencing, 48 (90.6%) were correctly detected by the CRDH. The first M. tuberculosis strain that was not correctly detected by the CRDH had a mutation at codon 513 (CAA'!CCA) and hybridized with all five WT probes, but the hybridization signal was weak at the site of the Rif1 probe. The second strain had a mutation at codon 516 (GAC'!TAC) and hybridized with all five WT probes, but the hybridization signal was weak at the site of the Rif2 probe. The third strain had point mutations at codons 511 (CTG'!CGC) and 516 (GAC'!GTC) and hybridized with all five WT probes, but the hybridization signal was weak at the site of the Rif1 and Rif2 probes, respectively. The fourth strain had point mutations at codons 516 (GAC'!GGC) and 529 (CGA'!CAA); the mutation at codon 516 was correctly detected by the lack of hybridization at the Rif2, but the mutation at 529 was not detected because of the presence of a hybridization signal at the Rif5. In the fifth strain with point mutations at codons 516 (GAC'!GTC) and 522 (TCG'!TTG), the assay correctly detected the mutation at codon 516 with no hybridization at the Rif2; however, the mutation at codon 522 was not detected due to the presence of a hybridization signal at the Rif3. All 103 strains without a mutation in the hot-spot region of the rpoB were hybridized correctly with all of the WT probes. The site of the mutation probes appeared clear while the spot of IS and CC were visualized as purple precipitates (C in Figure). The negative control PCR only bound to the spot of CC (B in Figure). When compared with the sequencing, the sensitivity and specificity of the CRDH were 90.6% and 100%, respectively. Analytical sensitivity and specificity of the CRDH - The sensitivity for rpoB and IS6110 sequences by PCR was 10 pg in agarose gel. The same amplicons were tested in the CRDH and the sensibility found was 1 pg (data not shown). The PCR products from the following strains did not hybridize with the probes: N. meningitidis, S. pneumoniae, S. enterica, H. influenzae, E. coli, M. marinum, M. intracellulare, M. scrofulaceum, M. gordonae, M. avium, M. smegmatis, M. kansasii, M. xenopi, M. fortuitum-peregrinum and M. phlei. GenoType MTBDR assay - The GenoType MTBDR results of 15 strains were compared with the CRDH. The same results were obtained by both the assays. Fourteen samples but one agreed with the sequencing results and had a correct detection of the 516 mutation, but the one strain, with point mutations at codons 516 and 529, only had a correct detection of the 516 mutation. Clinical isolates - The results of the CRDH for clinical isolates and the comparison with the AST and sequencing are shown in Table III. The CRDH was 100% concordant with the AST and sequencing. DISCUSSION The early identification of drug resistance would not only help to optimize the treatment of MDR-TB but also break the chains of transmission (Suresh et al. 2007). The CRDH assay can rapidly detect the presence of point mutations within the 81-bp of the rpoB gene and can identify the organism as a member of the MTBC. The test can successfully detect the five most common mutations of the rpoB gene. However, for isolates with other mutations, the CRDH indicates the presence of genetic alteration. The assay showed high analytical sensitivity and specificity. This technique had already been proved very promising in previous study in which the katG315 mutation was successfully characterized in 100% samples compared to the DNA sequencing (Verza et al. 2009). Recently, a novel automated molecular test, Xpert MTB/RIF, used a hemi-nested real-time reaction PCR assay to amplify M. tuberculosis specific sequence of the rpoB gene (Boehme et al. 2010, Helb et al. 2010). According to Boehme et al. (2010), the MTB/RIF test was specific in 99.2% patients without TB. Among culture-positive patients, the Xpert MTB/RIF test identified 98.2% patients with smear-positive TB and 72.5% with smear-negative TB. Other studies have also developed approaches which combine an initial real-time PCR with internal inhibition assessment and a pyrosequencing assay to detect the RIF resistance (Halse et al. 2010). The INNO-LiPA Rif.TB and Genotype MTBDR use reverse hybridization approach similar to our CRDH assay. The INNO-LiPA Rif.TB can detect resistance to the RIF (Morgan et al. 2005), while the GenoType MTBDR assay is able to detect resistance to both INH and RIF simultaneously (Albert et al. 2010, Anek-Vorapong et al. 2010). Although these tests have presented high sensitivity and specificity, their applications are very limited in the developing countries because the commercial kits are very expensive. The CRDH assay correctly identified 92.3% RIF-resistant strains when compared with the AST. Only one isolate, which was RIF-resistant by the AST, was considered a WT with the CRDH. This result implies that the CRDH method has limitations in specific situations. Some mutations are outside the detection range or in some cases no mutation exists in the hot-spot region of the rpoB gene even though the isolate is resistant to the RIF. In this case, other resistance mechanisms, such as a permeability barrier or drug efflux pumps probably exist (Bártfai et al. 2001, Heep et al. 2001, Caws et al. 2006). When compared with the DNA sequencing, the assay correctly identified 90.6% RIF-resistant strains. In detail, one isolate with the 529 (CGA'!CAA) mutation showed weak hybridization signal at the Rif4 and a strong one at the Rif5. This result was not entirely unexpected because codon 529 is located close to the 3' end of the Rif4 probe and 5' initial of the Rif5 probe. However, this mutation is probably not clinically relevant as it was detected in a strain harbouring the more frequent 516 mutation (GAC'!GGC). Another isolate with a mutation at codon 513 (CAA'!CCA) showed a weak hybridization signal at the Rif1, probably due to the high GC content at the opposite end of this probe. We postulate that the other three strains may have had a nonspecific hybridization signal because of mixtures of M. tuberculosis strains. The sequencing results for these isolates did not indicate a mixed population. However, another study that compared two commercially available DNA line probe assay observed that the strip tests are better at detecting mixed bacterial populations than the sequencing (Mäkinen et al. 2006). The problem encountered in the identification of the 529 (CGA'!CAA) mutation by the Genotype MTBDR was probably similar to our results, as described above. The main limitation of the CRDH assay is some weak signals that led to false susceptible results. Other probes and hybridization conditions need to be tested to overcome this limitation. The CRDH could also detect the rpoB mutation of M. tuberculosis in the clinical specimens. The results were concordant when compared with the AST and sequencing. However, the limitation of our study was a smaller number of positive clinical samples (10 out of 30) and only three had mutations. Therefore, a large number of samples should be analyzed for further conclusions. Comparing the results with the sequencing, the sensitivity and specificity of the CRDH were 90.6% and 100%, respectively. Therefore, the CRDH test is a promising tool for the early and rapid routine detection of frequent M. tuberculosis RIF-resistant genotypes in the mycobacterial laboratory. The CRDH could also be used for the molecular epidemiological studies to monitor the spread of a cluster of drug resistant strains within a community or institution. The genotypic test has several advantages over classical and other molecular detection methods. With this genotypic test RIF-resistance in M. tuberculosis isolates can be predicted within a day. Compared with the PCR-based sequencing and DNA microarray technique, our method can be performed without expensive devices, whereas the current commercial kits are of limited use in the laboratories of developing countries. However, the molecular drug susceptibility testing results must always be confirmed by phenotypic methods (Miotto et al. 2006, Hillemann et al. 2007). REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11023t3.jpg] [oc11023f1.jpg] [oc11023t2.jpg] [oc11023t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}