 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 2, 2011, pp. 146-152 ARTICLES The ability of haemolysins expressed by atypical enteropathogenic Escherichia coli to bind to extracellular matrix components Caroline A Magalhães; Sarita S Rossato; Ângela S Barbosa; Thiago O dos Santos; Waldir P Elias; Marcelo P Sircili; Roxane MF Piazza+ Laboratório de Bacteriologia, Instituto Butantan, São Paulo, SP, Brasil + Corresponding author: roxane@butantan.gov.br Received 6 August
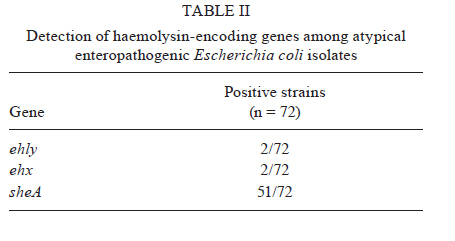
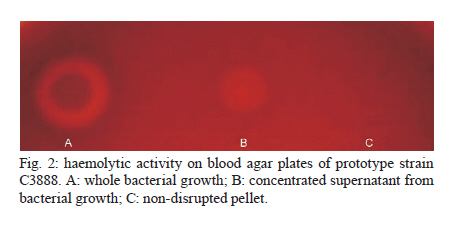
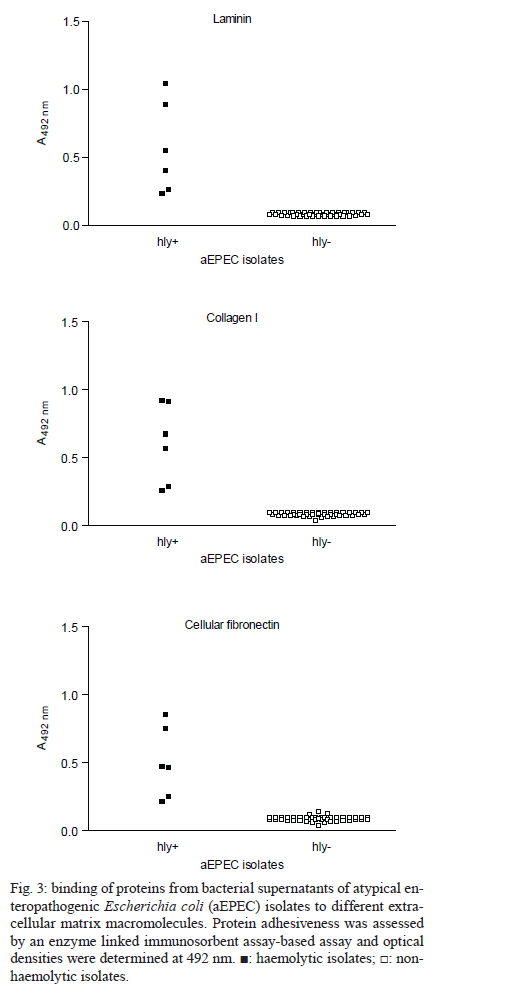
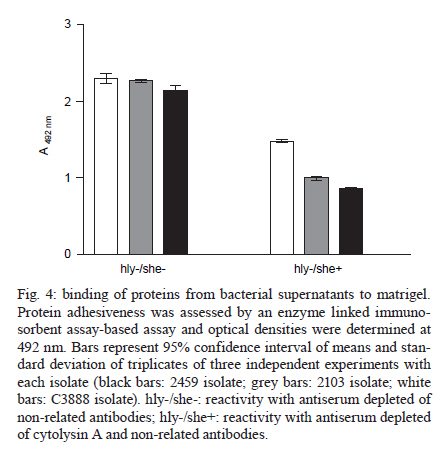
2010 Code Number: oc11024 ABSTRACT Typical and atypical enteropathogenic Escherichia coli (EPEC) are considered important bacterial causes of diarrhoea. Considering the repertoire of virulence genes, atypical EPEC (aEPEC) is a heterogeneous group, harbouring genes that are found in other diarrheagenic E. coli pathotypes, such as those encoding haemolysins. Haemolysins are cytolytic toxins that lyse host cells disrupting the function of the plasma membrane. In addition, these cytolysins mediate a connection to vascular tissue and/or blood components, such as plasma and cellular fibronectin. Therefore, we investigated the haemolytic activity of 72 aEPEC isolates and determined the correlation of this phenotype with the presence of genes encoding enterohaemolysins (Ehly) and cytolysin A (ClyA). In addition, the correlation between the expression of haemolysins and the ability of these secreted proteins to adhere to extracellular matrix (ECM) components was also assessed in this study. Our findings demonstrate that a subset of aEPEC presents haemolytic activity due to the expression of Ehlys and/or ClyA and that this activity is closely related to the ability of these isolates to bind to ECM components. Key words: enteropathogenic Escherichia coli - atypical EPEC - enterohaemolysins - cytolysin A - extracellular matrix components Enteropathogenic Escherichia coli (EPEC), one of the six diarrheagenic E. coli (DEC) pathotypes, expresses an adherence factor that is chromosomally encoded by the eae gene (EPEC attaching and effacing), located within the locus for enterocyte effacement (LEE) pathogenicity island (McDaniel & Kaper 1997). EPEC strains have been categorized into two subgroups, termed typical and atypical EPEC (aEPEC) (Kaper 1996). In addition to the LEE pathogenicity island, typical EPEC (tEPEC) strains contain Per regulators and the EPEC adherence factor plasmid (EAF), which encodes the bundle-forming pili (BFP) that mediate localized adherence to epithelial cells (Donnenberg et al. 1992). EPEC strains lacking the EAF plasmid (Kaper 1996) and that do not express BFP have been designated aEPEC (Abe et al. 2009, Hernandes et al. 2009, Nara et al. 2010). The aEPEC subgroup has been considered an emerging bacterial pathogen, associated with both sporadic cases and outbreaks of diarrhoea (Hernandes et al. 2009). Indeed, aEPEC isolates have been associated with diarrhoea in several countries, including Brazil (Afset et al. 2004, Robins-Browne et al. 2004, Franzolin et al. 2005, Nguyen et al. 2006, Bueris et al. 2007, Estrada-Garcia et al. 2009, Moreno et al. 2010). In Brazil, aEPEC is currently more prevalent than tEPEC, which was previously the most common bacterial agent in childhood endemic diarrhoea (Hernandes et al. 2009). These observations indicate that aEPEC constitutes an emerging DEC pathotype that requires epidemiological and virulence studies. Many bacterial pathogens produce toxins that lyse host cells by disrupting the function of the plasma membrane. The majority of these cytolytic toxins are pore-forming proteins and several of them are important virulence factors for their parent bacteria (Alouf 2001). In E. coli, several different pore-forming cytolysins have been identified. The most extensively studied is α-haemolysin (HlyA), which is produced by many uropathogenic E. coli strains. Its contribution to virulence has been shown in several animal models (Hull et al. 1981, Hacker et al. 1983). A toxin related to HlyA, enterohaemorrhagic E. coli (EHEC-HlyA, denoted Ehx), is encoded by ehx genes that are located on a virulence plasmid belonging to EHEC (Bauer & Welch 1996, Schmidt et al. 1996). Ehx has been identified in EHEC strains of serotype O157:H7 and is also present in most EHEC strains belonging to other serotypes, such as O26:H11/H- and O103:H2 (see Jenkins et al. 2008 for a review). In addition, it has been previously reported that EPEC O26:H- isolates are associated with a type of haemolysin, termed enterohaemolysin (Ehly) because of its association with enteric diseases, that is phenotypically, genetically and serologically unrelated to the previously described haemolysins (Beutin et al. 1989). Actually, the bacteriophage-associated proteins Ehly1 and Ehly2 in these isolates are not haemolysins per se. Instead, they induce the release of cytolysin A (ClyA) (Oscarsson et al. 2002a), a pore-forming haemolysin produced by E. coli K-12 that is unrelated to HlyA. ClyA is a 34-kDa protein that is encoded by a chromosomal gene named clyA (also referred to as hlyE and sheA) (Oscarsson et al. 1996, del Castillo et al. 1997). In addition to E. coli K-12, this haemolysin has also been shown to be produced by pathogenic E. coli, such as Shiga-toxin producing E. coli (STEC), enteroinvasive E. coli (EIEC), enteroaggregative E. coli (EAEC), enterotoxigenic E. coli (ETEC) (Ludwig et al. 1999) and extraintestinal E. coli isolates (ExPEC) (Kerényi et al. 2005). Haemolysins also play an important role in the pathogenesis of several other Gram-negative bacteria and they mediate adhesion to vascular tissue and/or blood components, such as plasma and cellular fibronectin (Lowrance et al. 1988). Therefore, in this study we investigated the expression of haemolysins, the presence of their encoding genes in aEPEC and the relationship between the expression and binding of these secreted proteins to extracellular matrix (ECM) macromolecules. MATERIALS AND METHODS Bacterial strains - Seventy-two aEPEC isolates were used in this study, all of which have been previously characterized by polymerase chain reaction (PCR) analysis as containing the eae gene and lacking the EAF plasmid, the bfpA gene and BFP expression (Bueris et al. 2007, Abe et al. 2009, Nara et al. 2010). These isolates belong to a wide range of serotypes and exhibit distinct patterns of adhesion to HEp-2 cells (Abe et al. 2009). Isolates C3888 (O26:H-) (Beutin et al. 1988) and BA86 (O76:H19) (Bueris et al. 2007) were used as prototypes for Ehly expressing and non-expressing isolates, respectively. Culture media and growth conditions - The following media were used: tryptic soy broth (TSB) (1.7% casein, 0.5% NaCl, 0.25% K2HPO4, 0.3% peptone and 0.25% glucose), Evans medium (EM) (0.15% yeast extract, 0.25% NaCl, 0.871% KH2PO4, 0.0005% MnCl2.4H2O, 0.005% MgSO4 and 0.0005% FeCl3.6H2O) and minimal medium (MM) (6.4% Na2HPO4.2H2O, 0.25% NaCl, 0.5% NH4Cl, 15% casein and 2% lactose). All reagents were purchased from Difco. Bacterial isolates were cultured in 3 mL of TSB, EM or MM with shaking at 150 rpm for 18 h at 37ºC. Stock cultures were made by taking aliquots corresponding to one optical density bacterial growth, supplementing them with glycerol to 80% (V/V) and storing them in cryotubes at -80ºC. Haemolytic activity on sheep blood agar plates - To detect haemolytic activity, bacteria were spotted onto blood agar plates directly from stock cultures or after pre-culturing in different liquid media (TSB, EM or MM). Two microliters of the growth suspension were spotted onto blood agar plates supplemented with 10 mM calcium chloride and containing 5% defibrinated sheep erythrocytes washed in phosphate buffer saline (PBS) pH 7.4. The plates were examined after 24 h of incubation at 37ºC for the presence of lysis zones around areas of bacterial growth (Beutin et al. 1989). Detection of ehxA, ehly and sheA genes - The 72 aEPEC isolates were tested for the presence of ehxA, ehlyA and sheA genes by PCR. All primers used in this study were based on conserved regions of these genes. The ehxA gene was amplified with forward (ATGACAGTAAATAAAATAAAGAAC) and reverse (TCAGACAGTTGTCGTTAAAGTTG) primers (Cookson et al. 2007) using the following amplification cycle: 30 x (95ºC/45 sec, 53ºC/30 sec, 72ºC/2 min), 72ºC/7 min. The ehlyA gene was amplified with forward (GCCGGGCACGTCTGTTC) and reverse (GCGCGGTGCCGTTTCCTG) primers based on the published sequence (GenBank accession X70047), using the following amplification cycle: 30 x (94ºC/45 sec, 64ºC/30 sec, 72ºC/2 min), 72ºC/8 min. The sheA gene was amplified with forward (GAGGCGAATGATTATGACTG) and reverse (ACTTCAGGTACCTCAAAGAG) primers (Kerényi et al. 2005) using the following amplification cycle: 30 x (94ºC/1 min, 58ºC/1 min, 72ºC/2 min), 72ºC/8 min. The amplified fragments were analyzed by electrophoresis in 0.8% agarose gels, resulting in a 2,997 bp fragment, a 712 bp fragment and a 920 bp fragment for ehxA, for ehly and for sheA genes, respectively. Anti-supernatant polyclonal antibody - The haemolytic phenotype was determined in both non-disrupted pellet and concentrated supernatant (50X) fractions of the C3888 strain. The supernatant was obtained by growing the C3888 isolate in 50 mL of TSB while shaking at 150 rpm for 18 h at 37ºC, followed by centrifugation at 10,000 g for 15 min. The supernatant was filtered through a 0.22 μm membrane and protein concentration was determined by a Bradford assay. A 65-day-old New Zealand rabbit was immunized intramuscularly with a lyophilized culture supernatant corresponding to 200 μg of protein, re-suspended in 1 mL of distilled water and emulsified with Montanide adjuvant (SEPPIC-France). A booster dose was given after 15 days. Serum samples to be used as a negative control in specific antibody evaluation were obtained just before the first immunization by the auricular-vein method. Serum samples were also obtained 15 days after the last antigen injection and were subsequently analyzed by enzyme linked immunosorbent assay (ELISA) using C3888 supernatant as antigen. To be depleted of non-related antibodies, the rabbit antiserum was adsorbed against the aEPEC isolate BA86 (the non-haemolytic control) or BA3160 (a cytolysin expressing isolate). Briefly, the bacterial pellet was resuspended in 0.01 M PBS pH 7.2 containing 1 mM phenylmethylsulphonyl fluoride and centrifuged again at 10,000 g for 15 min. The sample was kept at -20ºC until use. The adsorption procedure was performed by homogenizing the bacterial pellet with 1.5 mL of the antiserum in 2 mL microcentrifuge tubes. The sample was agitated for 2 h at room temperature and then incubated for 16-18 h at 4ºC. The mixture was then centrifuged at 5,000 g for 10 min and the pellet was discarded. This procedure was repeated twice after which the serum was filtered through a 0.22 μm membrane. The depleted antiserum was stored at -20ºC until use. ECM components binding assay - All macromolecules were purchased from Sigma Chemical Co Adhesion of proteins from bacterial supernatants to individual macromolecules of ECM was analyzed according to Barbosa et al. (2006). ELISA plate wells (Nunc-Immuno plate, Maxi Sorp) were coated with 1 μg of laminin, collagen type I and IV, cellular fibronectin, plasma fibronectin or matrigel in 100 μL of PBS for 2 h at 37ºC. The wells were washed three times with PBS 0.05% Tween 20 (PBST) and then blocked with 200 μL of 1% BSA for 1 h at 37ºC. Twenty micrograms of protein from the isolates' supernatants were added per well in 100 μL of PBS and proteins were allowed to bind to the different ECM substrates for 1.5 h at 37ºC. The plates were washed three times with PBST, then incubated with anti-supernatant serum diluted 1:5.000 for 1 h. After three washes with PBST, 100 μL per well of a 1:10.000 peroxidase-conjugated goat anti-rabbit immunoglobulin G in PBS was added and incubated for 1 h. All incubations were done at 37ºC. The wells were washed three times and o-phenylenediamine (0.04%) in citrate phosphate buffer (pH 5.0) plus 0.01% (w/v) H2O2 was added. The reaction was allowed to proceed for 15 min and was then terminated by the addition of 50 μL of 8 M H2SO4. To detect the bound proteins, the absorbance at 492 nm was measured using a microplate ELISA reader (Labsystems Uniscience, Multiskan EX). Statistical analysis - The ELISA absorbance (A492 nm) data from duplicates of three independent experiments were presented as mean and standard error and were analyzed using GraphPad Prism 5.00® software. Differences were considered significant when p < 0.05, by Student t test. RESULTS The phenotypic haemolytic profile of 72 isolates previously defined as aEPEC (Abe et al. 2009) was evaluated. To detect haemolytic activity, all bacteria were taken from stock cultures and spotted onto blood agar plates. Data regarding toxin expression or secretion are associated with the composition of the culture media in which the bacteria are grown (Vilhena-Costa et al. 2006, Rocha & Piazza 2007). We investigated whether the haemolytic profile of aEPEC isolates was dependent on media components. Therefore, the isolates were pre-cultured in different media (TSB, EM or MM) before being spotted onto blood agar plates. The isolates 2103 (O26:H11), 2459 (O26:H11), 2775 (O113:H19) and 3160 (O110:HNM) showed haemolytic activity in a narrow zone of haemolysis (Fig. 1A-D). The culture media had no influence on these four haemolytic isolates or the control strain (C3888) (Fig. 1E). In contrast, haemolytic activity was detected in two other strains, 2062 (O171:H48) and 4192 (O11:H25), but only after pre-incubation in MM (Fig. 1F-G, Table I). We next focused on correlating the haemolytic phenotype with the presence of the ehx, ehly and sheA genes. The presence of ehx and ehly was detected only in two isolates (2103 and 2459) (Tables I, II). In addition, the sheA gene was detected in 51 isolates, a prevalence of 70.8% (Table II). In the C3888 strain, all three genes were detected, confirming previous observations (Oscarsson et al. 2002a). As an experimental tool for further investigations, we produced a rabbit polyclonal antiserum against proteins from the concentrated supernatant of the prototype strain C3888, which produces Ehlys and ClyA. We used the supernatant to produce the antiserum because the haemolytic activity was detected in the supernatant fraction rather than in the non-disrupted pellet (Fig. 2). This observation differs from previous reports, which describe the lack of haemolytic activity in the culture supernatant of strain C3888 (Beutin et al. 1990, Figueiredo et al. 2003). However, these authors detected such activity in disrupted bacterial cells collected by centrifugation. Haemolytic activity was also detected in concentrated supernatants of isolates BA2103, BA2459, BA2775 and BA3160 (data not shown). Using this antiserum, which had been previously adsorbed with the non-haemolytic aEPEC control (strain 86), we investigated whether proteins from the supernatant of haemolysin-expressing and non-expressing aEPEC strains would interact with ECM macromolecules. Laminin-1, collagen type I, collagen type IV, cellular fibronectin and plasma fibronectin were immobilized on microdilution wells and protein attachment was assessed by an ELISA-based method. As can be seen in Fig. 3, proteins from isolates that exhibit the haemolytic phenotype bound to all tested ECM macromolecules. A statistical analysis demonstrated significant differences between the mean binding capacity of haemolytic and non-haemolytic isolates. The strains 2103 and 2459, in which the genes ehly/ehxA and sheA were detected, presented higher binding capacity (A492 nm c.a 1) either to laminin (p < 0.0001 R2 0.70) or to collagen I (p < 0.0001 R2 0.77) or to cellular fibronectin (p < 0.0001 R2 0.72) than the isolates which contain only sheA (2775 and 3160, A492 nm c.a 0.5). This higher binding capacity was also exhibited by the control strain C3888, which also contains the genes ehly and sheA. In addition, those isolates that showed haemolytic activity only after being pre-cultured in MM exhibited the lowest binding capacity to ECM molecules (2062 and 4192, A492 nm c.a 0.25) (Fig. 3). Furthermore, isolates that did not present haemolytic activity on blood agar plates did not display any binding to ECM components (Fig. 3). To check the differences between the binding capacity of isolates that express either Ehly and Ehx or ClyA, the antiserum was adsorbed with a ClyA expressing strain (3160). Both antisera were used to perform an ELISA on matrigel (a mixture of laminin, collagen and fibronectin) to compare the reactivity of the 2103, 2459 and C3888 isolates. The differences between the binding capacities of these isolates could be attributed to Ehly and/or Ehx. These data confirm the high ability of the strains that express Ehlys and ClyA to bind to ECM components, as verified by the statistical significance between the two experiments for each strain (2103: p = 0.01, R2 0.96), (2459: p = 0.0028, R2 0.99) and (C3888: p = 0.02, R2 0.94) (Fig. 4). DISCUSSION Recent reports on the aetiology of acute diarrhoea indicate that aEPEC is an emerging DEC pathotype (Hernandes et al. 2009), necessitating epidemiological and virulence studies. Additionally, many questions have arisen about the additional virulence factors expressed by aEPEC in comparison to tEPEC (Trabulsi et al. 2002, Hernandes et al. 2009). Among these factors are haemolysins, which are widespread among bacterial pathogens, including EHEC, a pathogen that is phylogenetically close to aEPEC (Trabulsi et al. 2002, Bando et al. 2009). In the present work, we assessed the haemolytic profile of aEPEC, which had been isolated in an epidemiological study of acute diarrhoea (Bueris et al. 2007). In six aEPEC isolates, the haemolytic phenotype was observed after precisely 24 h of incubation on blood agar plates, suggesting that this phenotype was due to Ehlys. However, only two of the isolates were positive for ehxA/ehlyA genes (BA2103 and BA2459), corroborating recent reports in which the plasmid-encoded Ehly (encoded by ehxA) of EHEC O157:H7 was significantly associated with diarrhoea (Afset et al. 2008, Scaletsky et al. 2009). The sheA gene has also been detected in pathogenic E. coli, such as STEC, EIEC, EAEC, ETEC (Ludwig et al. 1999) and ExPEC (Kerényi et al. 2005). We also found a high prevalence of the sheA gene in aEPEC, not correlated with the haemolytic activity. Usually ClyA, or "silent haemolysin", causes haemolysis under the following conditions: sheA is present in high-copy-number plasmids, certain sheA regulator genes (mprA and slyA) are overexpressed (del Castillo et al. 1997), the transcription of the chromosomal sheA is derepressed (Westermark et al. 2000) or other global transcription factors, such as fumarate and nitrate reduction and catabolite gene activator protein, are present (Ralph et al. 1998, Westermark et al. 2000, Wyborn et al. 2004). One of these conditions might explain the haemolytic phenotype of the two isolates in which only the sheA gene was present and haemolysis occurred independently of medium composition (BA2775 and BA3160), whereas in the two others (BA2062 and BA4192), the haemolytic activity was present only when they were previously cultivated in MM, which contains lactose. Because the sheA gene remains silent under the usual laboratory growth conditions (del Castillo et al. 1997), our data would imply that increased lactose concentrations promote an oxidative condition (Gottesman 2004), which would induce ClyA expression. Thus, the data presented here demonstrate that only some of the tested strains express ClyA, which in turn indicates that this toxin can play a role only in a subset of the pathogenic aEPEC. The main aspect examined in the present study was the capacity of proteins released by both haemolysin-expressing and non-expressing aEPEC strains to interact with ECM components. No specific binding to the target ECM macromolecules was detected with proteins released by strains that did not express haemolysin. In contrast, the ability to bind to ECM molecules by proteins produced by aEPEC strains that did express haemolysin could be categorized in three subsets: (i) high binding capacity shared by those strains in which the ehlyA, ehxA and sheA genes were present, (ii) medium binding capacity present in isolates harbouring the sheA gene and also showing haemolytic phenotype under the usual laboratory growth conditions and (iii) low binding capacity exhibited by the strains in which the gene sheA is present and the haemolytic phenotype was induced only after pre-cultivation in lactose-containing MM. The differences in binding capacity amongst the haemolytic isolates, which was detected when an antiserum depleted of SheA antibodies was used, suggest that the remaining binding activity to ECM observed in supernatants of isolates BA2103, BA2459 and C3888 can be attributed to EhxA and/or Ehly activity. Further experiments, currently under way in our laboratory, will clarify the role of these haemolysins. Although cytolytic toxins are considered to be virulence factors and are found in disease-causing bacteria, their precise contribution to pathogenesis is not completely understood. In some Salmonella enterica serovars, including Salmonella Typhi, HlyE (ClyA or SheA) (Fuentes et al. 2008), was described as a pore-forming haemolysin that accumulates in the S. Typhi periplasm and can be artificially released by destabilizing the bacterial envelope (Oscarsson et al. 2002b, Fuentes et al. 2008). Purified E. coli HlyE is cytotoxic to mammalian cells and the functional transfer of S. Typhi HlyE to Salmonella typhimurium promotes deep organ infection in mice (Lai et al. 2000, Oscarsson et al. 2002b, Fuentes et al. 2008). The capacity of proteins secreted by haemolysin-expressing aEPEC isolates to bind to ECM, described here for the first time, represents one more piece in the complicated pathogenesis mechanism of aEPEC. These data suggest that the adhesion of haemolysins to matrix macromolecules might contribute to a second step in this process, as already described for other pathogens (Matsunaga et al. 2003, Liang & Ji 2006). In conclusion, the data presented in this study demonstrate that a subset of aEPEC presents haemolytic activity due to Ehly and/or ClyA. This activity involves the binding of proteins released by these isolates to ECM components, suggesting that these haemolysins can play a role in pathogenesis. ACKNOWLEDGEMENTS To Dr Claudia Trigo Pedroso de Moraes, for technical support. REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11024t1.jpg] [oc11024f3.jpg] [oc11024f2.jpg] [oc11024t2.jpg] [oc11024f4.jpg] [oc11024f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}