 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 2, 2011, pp. 153-157 ARTICLES Schistosoma mansoni: a method for inducing resistance to praziquantel using infected Biomphalaria glabrata snails Flávia FB CoutoI; Paulo Marcos Z CoelhoI, +; Neusa AraújoI; John R KuselII; Naftale KatzI; Liana K Jannotti-PassosIII; Ana Carolina A MattosI ILaboratório
de Esquistossomose, Instituto de Pesquisas René Rachou-Fiocruz, Belo
Horizonte, MG, Brasil + Corresponding author: coelhopm@cpqrr.fiocruz.br Received 9 August
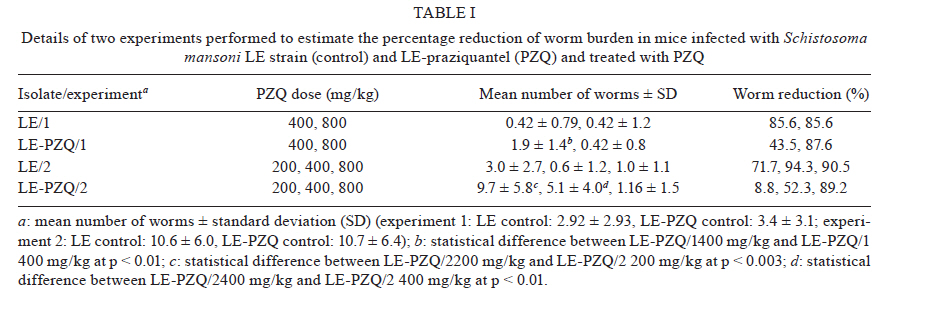
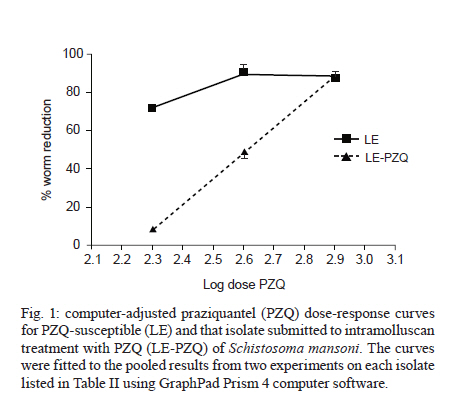
2010 Code Number: oc11025 ABSTRACT To elucidate the mechanisms of antischistosoma resistance, drug-resistant Schistosoma mansoni laboratory isolates are essential. We developed a new method for inducing resistance to praziquantel (PZQ) using successive drug treatments of Biomphalaria glabrata snails infected with S. mansoni. Infected B. glabrata were treated three times with 100 mg/kg PZQ for five consecutive days with a one-week interval between them. After the treatment, the cercariae (LE-PZQ) produced from these snails and the LE strains (susceptible) were used to infect mice. Forty-five days after infection, mice were treated with 200, 400 or 800 mg/kg PZQ. Thirty days post-treatment, we observed that the mean number of worms recovered by perfusion was significantly higher in the group of mice infected with the LE-PZQ isolate treated with 200 and 400 mg/kg in comparison to the LE strain with the same treatment. Moreover, there was a significant difference between the ED50 (effective dose required to kill 50% of the worms) of the LE-PZQ isolate (362 mg/kg) and the LE strain (68 mg/kg). In the in vitro assays, the worms of the LE-PZQ isolate were also less susceptible to PZQ. Thus, the use of infected snails as an experimental model for development of resistance to S. mansoni is effective, fast, simple and cheap. Key words: Schistosoma mansoni - Biomphalaria glabrata - praziquantel - resistance induction - new method The current strategy for schistosomiasis control is based on large-scale treatments of populations aimed at reducing disease morbidity (WHO 2002). Currently, praziquantel (PZQ) is the drug of choice (Utzinger & Keiser 2004, Fenwick & Webster 2006), with the main advantages of its use being oral administration, single dose, low toxicity and low cost (Fenwick et al. 2003, Utzinger & Keiser 2004). Despite the advantages of PZQ, there is concern about the development of Schistosoma mansoni resistance to PZQ, both under laboratory and field conditions (Abdul-Ghani et al. 2009). In the laboratory, induction of resistance is based on the treatment of mice infected with S. mansoni, initially using sub-curative doses of PZQ. Afterwards, the dosage is increased for at least seven passages in mice/snails to complete the life cycle of the parasite (Ismail et al. 1994, Fallon et al. 1995). The complete mechanism of action of PZQ is still unclear (Doenhoff et al. 2008). Obtaining resistant strains is important for the evaluation of such mechanisms as well as for the development of alternative drugs for schistosomiasis treatment and control. Studies show that PZQ is effective not only in adult worms, but also in the intramolluscan phase of the parasite (Coelho et al. 1988, Mattos et al. 2006, 2007). We report a novel method for the induction (or selection) of S. mansoni worms resistant to PZQ using successive treatments of infected Biomphalaria glabrata snails. SUBJECTS, MATERIALS AND METHODS Parasites and hosts - The S. mansoni (LE strain) life cycle was maintained using B. glabrata (Barreiro de Cima strain) snails as intermediate hosts and Swiss mice as definitive hosts, according to Pellegrino and Katz (1968) and Souza et al. (1995). Perfusion of adult worms from infected mice - Two methods were used. The methodology described by Pellegrino and Siqueira (1956) used a needle attached to a Brewer's automatic pipetter to inject saline solution under pressure into the descendent aorta. Afterwards, saline was injected into the hepatic hilum of mice after sectioning the portal vein, allowing the perfusion of the portal system and mesenteric veins. Worms were recovered and counted. To recover the worms using the methodology described by Smithers and Terry (1965) the portal vein of the mice was sectioned and the culture medium was gently injected into the base of the left ventricle of the infected mice's hearts. It is not possible to recover all the worms using this methodology with a lower pressure injection, but the integrity of the parasite's tegument is preserved. Therefore, this methodology is ideal for the recovery of worms when the goal is to cultivate or evaluate other parameters such as tegumental integrity and/or excretory activity. Induction of resistance to PZQ in the intramolluscan phase - Two-hundred B. glabrata were individually infected with 10 S. mansoni miracidia (Mattos et al. 2007). Thirty days post-infection the snails were exposed to artificial light for 1 h to verify the shedding of cercariae (Souza et al. 1995). The snails shedding cercariae were divided into two groups (infected snail or infected snails treated with PZQ). The snails were individually weighed using an analytical balance and the snails' mean weight was used to calculate the drug dosage to be administered. The snails were individually treated with 100 mg/kg PZQ for five consecutive days. This treatment was repeated three times, with a one-week interval between the treatments. PZQ was added to the 100 mg of food (mouse chow; Nuvital) daily allowance. The ration was reconstituted in water until it became pasty and then PZQ was added. The untreated snails received the same amount of food daily without PZQ (Mattos et al. 2007). The daily amount of food given each day was totally ingested by each snail, suggesting that the snails received all of the drugs offered in ration. The snails were kept in plastic glasses containing 200 mL dechlorinated water which was changed daily. At the end of the treatments, the snails were exposed each week to artificial light until cercarial shedding occurred in sufficient number for the infection of mice, which occurred in about 60 days. Evaluation of PZQ pressure on parasites at the intramolluscan phase - Treatment with PZQ: mice were infected with 25 S. mansoni cercariae by the subcutaneous route (Pellegrino & Katz 1968). Eighty-four mice were infected with cercariae derived from snails treated with PZQ, called the LE-PZQ isolate and another 84 mice infected with cercariae developed from untreated snails, called the LE strain. Forty-five days after infection, mice were divided into groups (12 mice in each group). Two experiments were performed: in the first the mice were treated with 200 or 400 mg/kg PZQ; in the second experiment, mice were treated with 200, 400 or 800 mg/kg. In both experiments a single oral dose was used. An untreated group was included as a control. Thirty-three days after treatment, mice were submitted to perfusion for evaluation of worm burden reduction (Pellegrino & Siqueira 1956). The percentage of reduction for each group and each experiment was calculated according to the following formula:
The mean of worm reduction related to the experiments 1 and 2 for each PZQ dose was calculated and then the effective dose required to kill 50% of the worms (ED50) was estimated. Data were analyzed using Minitab 13 and GraphPad Prism 4 computer software using the sigmoidal curve, with 5% significance. In vitro observation of worms treated with PZQ - Mice infected with 100 cercariae derived from S. mansoni LE or LE-PZQ isolates were treated with 400 mg/kg PZQ 45 days post-infection. Two hours later, mice were perfused using Roswell Park Memorial Institute medium (RPMI) 1640 medium with 2% heparin (Smithers & Terry 1965). Four pairs of worms originating from LE or LE-PZQ isolates were put in 6-well culture plates containing 4 mL RPMI 1640 supplemented with 5% foetal bovine serum and 100 μg/mL of penicillin and streptomycin, and were cultivated in a 5% CO2 incubator for eight days at a temperature of 37ºC. The worms derived from LE or LE-PZQ isolates and recovered from untreated mice were also cultivated and used as a control. RESULTS Evaluation of PZQ pressure in parasites at the intramolluscan phase - Treatment with PZQ - in this experiment, two doses of PZQ (400 or 800 mg/kg) were tested on mice infected with LE and LE-PZQ isolates. A statistically significant difference (p < 0.03) was observed between the mean number of worms recovered from LE-PZQ (1.9) and the mean from the LE strain (0.42) when both were exposed to 400 mg/kg PZQ. Moreover, the percentage reduction of worms from the LE-PZQ (43.5%) isolate was lower than that found for worms from the LE strain (85.6%), suggesting that the LE-PZQ isolate showed a reduction of susceptibility to drug action. When the higher dose of 800 mg/kg was used, no significant difference could be observed between the isolates (Table I). In the second experiment, three doses of PZQ (200, 400 or 800 mg/kg) were used. The mean number of worms recovered from the LE-PZQ isolate treated with 200 mg/kg (mean of 9.7) and treated with 400 mg/kg (mean of 5.1) was significantly lower (p < 0.002 and p < 0.01, respectively) than the mean of worms recovered from the groups infected with cercariae derived from the LE strain and treated with the same doses (mean of 3.0 and 0.6, respectively). Moreover, the percentage in reduction of worms was as follows: LE-PZQ treated with 200 mg/kg, 8.8%, with 400 mg/kg, 52.3%, LE treated with 200 mg/kg, 71.7%, and with 400 mg/kg, 94.3%. No statistically significant difference between the isolates could be detected when they were submitted at the dose of 800 mg/kg (Table I), corroborating our previous results. The mean worm reduction in experiments 1 and 2 was calculated for each PZQ dose (200, 400 or 800 mg/kg) (Fig. 1, Table II). Based on these means, the ED50 values obtained were 68 mg/kg and 362 mg/kg for the LE and LE-PZQ isolates, respectively, with these values being significantly different (p < 0.01) (Table II). The ED50 for the LE-PZQ isolate was 5.3 times higher than that for the LE strain. These results show that there is a reduction in S. mansoni susceptibility to PZQ when the parasite is submitted to PZQ treatment at the intramolluscan phase. In vitro observation of worms recovered from mice treated with PZQ - It was observed that the parasites of the LE strain derived from mice treated with 400 mg/kg PZQ were more contracted (Fig. 2A) than the parasites of LE-PZQ isolate submitted to the same PZQ dose (Fig. 2B). Additionally, more than 300 eggs were found in the cultured LE-PZQ isolate recovered from mice treated with 400 mg/kg. Furthermore, eggs in all stages were observed after eight days (Fig. 2D). In contrast, the worm culture of the LE strain had a smaller number of dead eggs (± 30 from total number of cultured worms) (Fig. 2C). The untreated LE and LE-PZQ isolates showed a similar number of eggs from the first stage to the mature stage (Fig. 2E, F). These results suggest that LE-PZQ isolates have reduced susceptibility to the drug. DISCUSSION According to Roos et al. (1993) and Liang et al. (2003), the process of drug pressure for selection of resistance can be carried out in the laboratory. This is important since resistant parasites may help elucidate the mechanism of the drugs. The PZQ effective against only adult worms and another drugs effective against adult and immature S. mansoni worms in the vertebrate host are also effective at the intramolluscan stage of the parasite, leading to interruption of shedding of cercariae (Warren & Weisberger 1966, Warren 1967, Coles 1979, Touassem & Combes 1986, Coles & Bruce 1987, Yi & Combes 1987, Coelho et al. 1988, Riley & Chappell 1990, Mattos et al. 2006, 2007). Mattos et al. (2007) tested different doses and schedules of PZQ in infected B. glabrata and observed that a treatment of 1,000 mg/kg PZQ significantly interrupted almost 90% of the cercarial shedding until approximately 30 days after the end of treatment, whereas treatments with other drug formulations (500 mg/kg PZQ and 250 mg/kg oxamniquine in association) eliminated the parasite from the snails. Thus, the dose of 100 mg/day/snail used in this study represents a dose lower than that required to eliminate the parasite. Furthermore, three treatments were carried out for five consecutive days with the goal being to expose the parasite periodically to PZQ to select or induce resistance. The results presented here demonstrate that adult worms of the LE-PZQ isolate were less susceptible to PZQ at doses of 200 and 400 mg/kg than the control strain (LE). Similar results were not observed between the LE and LE-PZQ isolates at a dose of 800 mg/kg. This dose was deemed to be enough to overcome the resistance of the selected parasite. Interestingly, a dose of 800 mg/kg in the susceptible strain (LE) shows the same results as with 400 mg/kg. However, when the dose is increased to 800 mg/kg, the drug kills around 88% of worms in the resistant isolate (LE-PZQ). In our in vitro experiments, adult worms obtained from mice treated with PZQ from the LE-PZQ isolate were less susceptible to the drug. The LE-PZQ isolate has been maintained in our laboratory under the pressure of PZQ. Recent results confirm that this isolate remains less susceptible to PZQ (unpublished data). The complete mechanism of action of PZQ has not been clearly defined (Doenhoff et al. 2008), but it is well known that PZQ causes a severe muscular contraction due to intense calcium influx (Pax et al. 1978), immediate tegumental lesions (Harnett & Kusel 1986), reduction of glutathione levels (Ribeiro et al. 1998) and interruption of the excretory activity of adult worms (Kusel et al. 2006, 2007, Oliveira et al. 2006). After eight days of culture the adult worms of the LE strain showed typical alterations in morphology of susceptible parasites in contact with the drug including intense contraction, reduced oviposition and tegumental changes. Alternatively, PZQ (at 400 mg/Kg) was not able to cause the same alterations in the motility and oviposition in the worms from the LE-PZQ isolate. The results obtained in the in vitro experiment are significant because they allow us to observe the worm condition immediately after contact with PZQ. We found that the ED50 for the LE-PZQ isolate was significantly higher than for the LE strain in both experiments. This parameter has been used to compare susceptible and resistant worms. Cioli et al. (2004) estimated the ED50 from four susceptible and five putatively PZQ-resistant isolates and suggested that the value of ED50 is useful for establishing and monitoring the drug susceptibility/resistance profiles of the parasite strain. Further, they observed that the ED50 value was three times higher in the resistant isolates when compared with the susceptible ones. Our results demonstrate that the ED50 of the LE-PZQ isolate was 5.3 times higher in comparison with the ED50 for the LE strain (susceptible). Both Fallon et al. (1996) and Bennett et al. (1997) suggested that the ED50 for resistant isolates should be at least five times higher than that for those isolates susceptible to the drug. For Bennett et al. (1997), a three-to-five times higher dose is sufficient to differentiate between susceptible and resistance strains. According to Fallon et al. (1996), induction of resistance performed in the laboratory using successive passages and treatments in mice are time-consuming and use dissimilar methodologies, thus rendering a difficult comparison of the analysis of the data. The study by Fallon and Doenhoff (1994) aimed at selecting PZQ resistant S. mansoni isolates was carried out with several passages of using the entire life cycle in mice and snails, with the infected mice being submitted to treatments with increasing doses of PZQ. This model for selection of resistant S. mansoni lineage is complex and time-consuming, but is able to select parasites at the adult worm phase. In our experimental studies, the chemotherapeutic pressure occurs on a higher number of parasites (thousands of cercariae and sporocysts) in the snail intermediate host. Thus, the selection of resistant strains in the laboratory using S. mansoni at the intramolluscan phase seems to be more useful and this approach may be very important in studies concerning the development of novel antischistosomal drugs, as well as in those aimed at gaining a better understanding of the mechanisms related to resistance. These experiments must be repeated with other isolates. In conclusion, the use of the snail as an experimental model for development of resistance in S. mansoni is an effective, fast, simple and cheap method. ACKNOWLEDGEMENTS To Vera de Paula Ribeiro, for translating our manuscript, and to the technicians of the Mollusc Room and the Instituto de Pesquisa René Rachou-Fiocruz, for maintenance of the parasite life cycle. REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11025f1.jpg] [oc11025t2.jpg] [oc11025t1.jpg] [oc11025f2.jpg] |
| |||||||||


{kind=link}
{kind=link}
{kind=link}
{kind=link}