 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 2, 2011, pp. 158-165 ARTICLES Studying nanotoxic effects of CdTe quantum dots in Trypanosoma cruzi Cecilia Stahl VieiraI; Diogo Burigo AlmeidaII; André Alexandre de ThomazII; Rubem Figueredo Sadok Menna-BarretoIII; Jacenir Reis dos Santos-MalletI; Carlos Lenz CesarII; Suzete Araujo Oliveira GomesI, IV; Denise FederIV, + ILaboratório
de Transmissores de Leishmanioses, Setor de Entomologia Médica e Forense + Corresponding author: mdfeder@id.uff.br Received 20 August
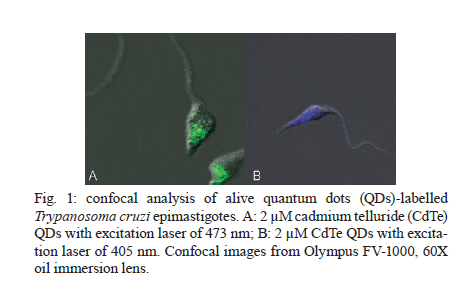
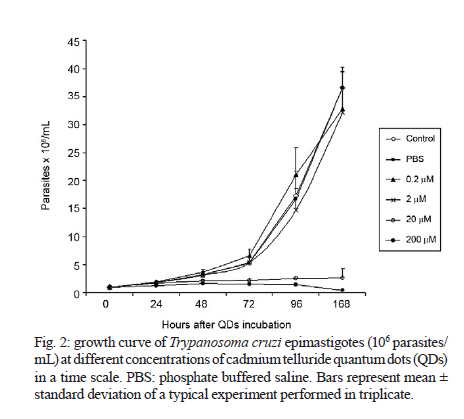
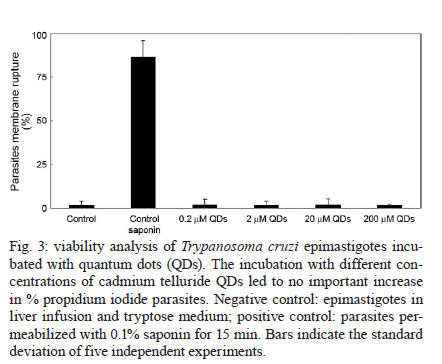
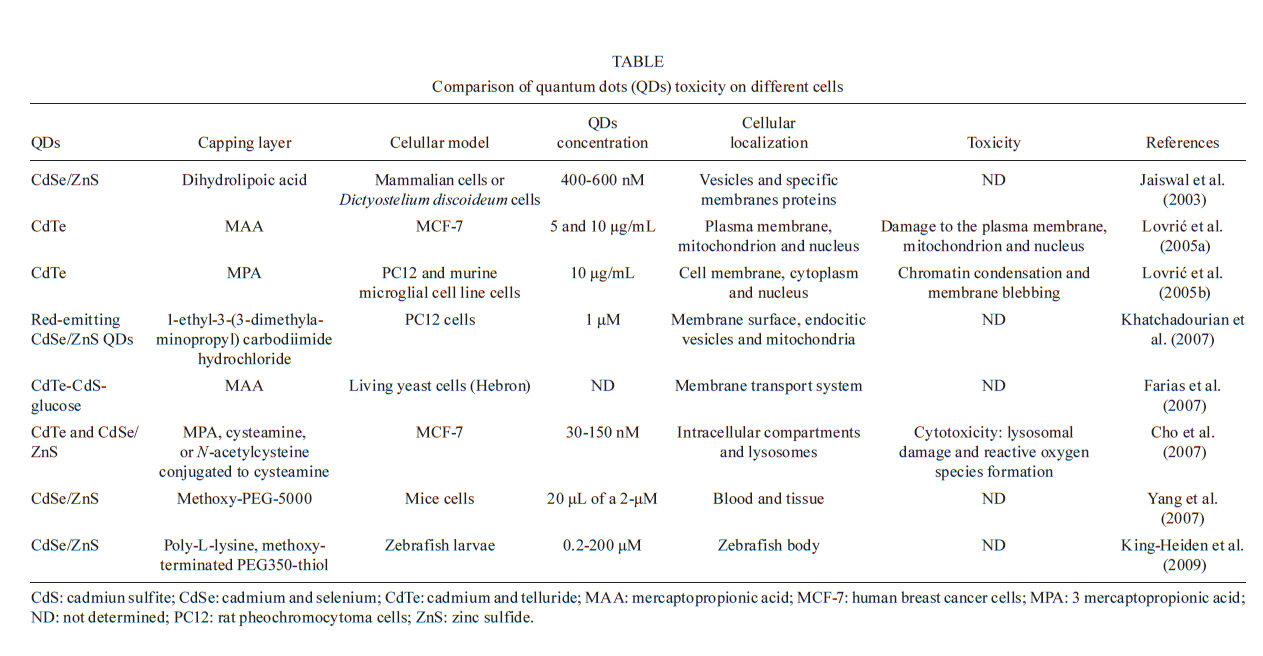
2010 Code Number: oc11026 ABSTRACT Semiconductor nanoparticles, such as quantum dots (QDs), were used to carry out experiments in vivo and ex vivo with Trypanosoma cruzi. However, questions have been raised regarding the nanotoxicity of QDs in living cells, microorganisms, tissues and whole animals. The objective of this paper was to conduct a QD nanotoxicity study on living T. cruzi protozoa using analytical methods. This was accomplished using in vitro experiments to test the interference of the QDs on parasite development, morphology and viability. Our results show that after 72 h, a 200 μM cadmium telluride (CdTe) QD solution induced important morphological alterations in T. cruzi, such as DNA damage, plasma membrane blebbing and mitochondrial swelling. Flow cytometry assays showed no damage to the plasma membrane when incubated with 200 μM CdTe QDs for up to 72 h (propidium iodide cells), giving no evidence of classical necrosis. Parasites incubated with 2 μM CdTe QDs still proliferated after seven days. In summary, a low concentration of CdTe QDs (2 μM) is optimal for bioimaging, whereas a high concentration (200 μM CdTe) could be toxic to cells. Taken together, our data indicate that 2 μM QD can be used for the successful long-term study of the parasite-vector interaction in real time. Key words: CdTe quantum dots - Trypanosoma cruzi - nanotoxicity Trypanosoma cruzi (family Trypanosomatidae, order Kinetoplastida), the etiologic agent of Chagas disease (Chagas 1909), is transmitted to humans by either blood sucking triatomine insect vectors, blood transfusion, organ transplantation or congenital transmission. Chagas disease is present in 18 tropical and subtropical countries and has affected 17 million individuals (Moncayo & Silveira 2009). Various fluorescent methods have been developed and utilized for T. cruzi studies. Previous investigations have demonstrated that normal biological function is maintained in the long term by living protozoan parasites such as T. cruzi after labelling with cadmium telluride (CdTe) quantum dots (QDs) (Feder et al. 2009). Therefore, QDs are considered to be safe biomarkers to study the parasite-vector interaction (T. cruzi-Rhodnius prolixus). However, detailed investigations of the toxicity of QDs in parasites or parasite-cell interactions are still scarce. Nanotoxicology is an emerging discipline that attempts to characterize and categorize the effects of engineered nanomaterials and to determine relationships between nanoparticles and toxicity (Marquis et al. 2009) that can be used to formulate rules for the design of safer nanomaterials. QDs are semiconductor nanocrystals used in the electronics industry and biomedical imaging. The main advantage of QDs in biomedical imaging is their high photostability, which allows long-term real-time study of in vitro, in vivo and ex vivo cell processes (Feder et al. 2009, Kasemets et al. 2009, Marquis et al. 2009). However, this advantage would be overridden if QDs' toxicity to cells can change their functionality and cause artefacts. For example, Cadmium (Cd) and Tellurium (Te), two widely used constituent metals in QD core metalloid complexes, are known to cause acute and chronic toxicity in vertebrates (Kondoh et al. 2002, Ha-milton 2004). Cd, a probable carcinogen, has a biological half-life of 15-20 years in humans, bioaccumulates, is known to cross the blood-brain barrier and placenta, and is systemically distributed to all bodily tissues, with liver and kidney being target organs of toxicity (Hardman 2006). If Cd atoms dissociate from QDs, they would be toxic to the organism. QDs can be coated with inert materials, such as silica, to prevent dissociation, to make QDs less toxic than conventional organic dyes and to create electron-dense structures (Hoshino et al. 2004). Nanoparticle toxicity is determined by its individual physicochemical characteristics (Hardman 2006). Size, charge, concentration, outer coating bioactivity (capping material and functional groups) and oxidative, photolytic and mechanical stability can determine QDs' toxicity both collectively and individually. Studies specifically designed for toxicologic assessment (e.g., dose, duration, frequency of exposure and mechanisms of action) are rare, but some reports have indicated a dose-dependent cytotoxicity (Hoshino et al. 2004). Several studies have suggested that the cytotoxic mechanism of QDs involve photolysis or oxidation. Under oxidative and photolytic conditions, the coatings of QDs are labile and are subject to degradation, which exposes potentially toxic "capping" material or intact core metalloid complexes or results in the dissolution of the core complex into the QDs' core metal components [e.g., Cd, Selenium (Se)] (Derfus et al. 2004, Kirchner et al. 2005, Hardman 2006). It had been suggested that certain types of QDs may be cytotoxic; CdTe QDs coated with mercaptopropionic acid and cysteamine were cytotoxic to rat pheochromocytoma (PC12) cell cultures at concentrations of 10 μg/mL. Uncoated CdTe QDs were cytotoxic at 1 μg/mL (Lovri et al. 2005a, Hardman 2006). Previous reports have described the cytotoxicity of CdTe QDs in mammals, bacteria and protozoan cells (Jaiswal et al. 2003, Dumas et al. 2009). Some of the observed cellular changes suggest necrotic or apoptotic phenotypes characterized by chromatin condensation and membrane blebbing. Moreover, the major mechanism for QD toxicity has been determined to be oxidative stress or DNA damage (Lovri et al. 2005a, Cho et al. 2007). DNA damage may be associated with apoptosis and oxidative stress because the nanoparticles are in direct contact with the cell nucleus (Hardman 2006, Marquis et al. 2009). However, previous studies have clearly showed that cells containing QDs could maintain cell function. Cells labelled with QDs remained viable for study over several days and generations (Parak et al. 2005). In one study, mammalian (HeLa) cells or Dictyostelium discoideum cells did not present any alterations and continued the endocytic process of QDs after 20 days (Jaiswal et al. 2003). Similar experiments performed by our group have demonstrated that insects fed QDs presented no significant apparent dysfunction or toxicity for up to 10 days (Feder et al. 2009). In this context, the goal of the current study was to evaluate the toxicity of our CdTe QDs on T. cruzi as a function of QD concentration and time exposure and to investigate the parasite's viability and nanoparticle localization. MATERIALS AND METHODS Parasites - T. cruzi epimastigotes (Dm28 clone), classified as TcI (Anonymous 1999), were grown at 28ºC in liver infusion and tryptose with hemin and folic acid supplemented with 10% bovine foetal serum (Chiari & Camargo 1984, Jaffe et al. 1984). All parasites were collected during an exponential growth phase. QDs preparation - To obtain a green-emitting sample of QDs, we diluted 3.8 mg of Te powder (approximately 40 mesh) in 40 mL of Milli-Q water. This solution was placed in a three-neck flask under argon atmosphere and stirred vigorously for 15 min to purge dissolved oxygen. Next, 35 mg of NaBH4 in 1 mL of water was added to the solution to reduce metallic Te to Te2-. The Te reduction process was performed at 80ºC for approximately 2 h. A secondary solution was produced by mixing 0.52 mL of a 0.1 mol/L solution of Cd perchlorate with 0.24 mL of a 4.7% of mercaptoacetic acid (MAA) solution. The pH of this secondary solution was adjusted to 11 and injected into the primary solution resulting in a colloidal CdTe QD solution. CdTe QDs were built from the adaptation of two methods: Zhang et al. (2000) and Gaponik et al. (2002). For details, refer to the Materials and Methods section in the study by Feder et al. (2009). Parasites labelling - For the in vitro assay, 5 μL of a T. cruzi (1X107) suspension was incubated with 50 μL of diluted yellow-emitting CdSe QDs (10 μL CdSe to 90 μL of a 0.14 M PBS/0.01 M NaCl phosphate buffer, pH 7.2) and stabilized with MAA, for 60 min at 28°C (Feder et al. 2009). The parasites were then observed by confocal microscopy. Cell proliferation - To determine the effect of QDs on live parasite cells, we constructed a dose-response lethality curve using several concentrations. On the fifth day of culture, parasites were harvested in sterile tubes, centrifuged at 1,500 g at 4ºC for 10 min and washed twice with the cold buffer solution [phosphate buffered saline (PBS)] described above. The cell density was estimated using a haemocytometric chamber and the growth curve was initiated with 1 x 106 cells followed by seven days of incubation with different CdTe concentrations: 0.2, 2, 20 and 200 μM. Cell proliferation was verified at 0, 24, 48, 72, 96 and 168 h after incubation with the nanoparticles. Flow cytometry - For flow cytometry analysis, parasites (3 x 106 cells in 300 μL) were incubated with QDs for 72 h. To evaluate cell viability, parasites were incubated with 30 μg/mL of propidium iodide (PI) (Sigma-Aldrich, Saint Louis, USA) for 15 min at 28ºC. For cell-cycle analysis, parasites were permeabilized with 0.1% saponin for 15 min before incubation with 30 μg/mL PI to improve PI labelling (Menna-Barreto et al. 2007). Data collection and the analysis were conducted using the FACScalibur flow cytometer (Becton-Dickinson, San Jose, USA). A total of 10,000 events were acquired in the regions previously established for T. cruzi epimastigotes. The comparison between control and QD groups were made using the Mann-Whitney test. Differences were considered statistically significant with p < 0.05. Transmission electron microscopy - Epimastigotes incubated with different concentrations of CdTe QDs were fixed in 2.5% glutaraldehyde diluted in a 0.1 M cacodylate buffer, pH 7.2, for 2 h at room temperature and postfixed in a 0.1 M cacodylate buffer containing 1% OsO4, 5 mM calcium chloride and 0.8% potassium ferricyanide for 1 h. The cells were dehydrated in a graded series of acetone and embedded in epoxy resin. Ultrathin sections were stained with uranyl acetate and lead citrate for observation under a Jeol JEM 1011 transmission electron microscope (Tokyo, Japan). RESULTS Live parasite cell imaging following incubation with CdTe QDs - Our previous studies demonstrated that parasites labelled with green-emitting CdTe QDs showed a strong, unspecific fluorescence and maintained cell integrity and intense movements (see movies at Feder 2009a, b). Parasites labelled with 2 μM of QDs are included as a reference (Fig. 1). QD dose-response curve in live parasites - Based on a time-scale growth curve, we determined the doses of CdTe for which T. cruzi maintained normal (i) integrity for at least seven days, (ii) proliferation and (iii) motility. The results showed that the effects are dose- dependent but not time-dependent. Parasites incubated with 20 and 200 μM CdTe QDs could not proliferate at similar rates as the control. Fig. 2 shows the exponential growth curve of these parasites over time, with no significant differences between the control, 2 and 0.2 μM of CdTe groups until after 168 h (7 days). Treatments with 20 and 200 μM hindered the growth of parasites when compared with the control group. The integrity and motility of these parasites remained intact up to 96 h. Cellular viability was assessed before and after incubation using the motility and Trypan blue cell dye exclusion methods (Gomes et al. 2006). Fig. 2 also shows that after 72 h, the 0.2 and 2.0 μM groups exhibit significant differences in growth. A significant increase in parasite death was observed for the 20 and 200 μM CdTe QD groups. Viability of T. cruzi epimastigotes - Parasite viability was checked after 72 h of incubation. The negative control consisted of parasites without QDs and PI. The positive control consisted of parasites that were permeabilised with 0.1% saponin and PI, as described in Materials and Methods. After 72 h, the incubation with QDs (0.2-200 μM) did not cause plasma membrane disruption. The positive control with saponin-permeabilized parasites showed a high percentage of PI+ cells (about 75%) (Fig. 3). Cell cycle of T. cruzi epimastigotes - The genetic content of T. cruzi was evaluated by flow cytometry via observation of the following: (i) the percentage of fragmented DNA, (ii) interphasic DNA, which indicates that the parasites are not dividing and (iii) duplicated DNA, which indicates that the parasites are undergoing mitosis. According to our results shown in Fig. 4, we observed a significant increase in DNA fragmentation with CdTe QDs that was dose-dependent in parasites incubated with 20 and 200 μM (p < 0.011 and p < 0.006, respectively). No significant differences were observed for interphasic DNA. However, the incubation with 2, 20 and 200 μM of CdTe QDs showed less duplicated DNA, indicating at least a partial arrest of T. cruzi mitosis (p < 0.015; p < 0.012 and p < 0.004, respectively). Transmission electron microscopy - Ultrastructural analysis of epimastigotes incubated with 2 μM CdTe QDs showed the integrity of cellular organelles such as mitochondria, nucleus, Golgi complex and plasma membrane (Fig. 5A, B). We observed QDs in vesicles scattered in the cytoplasm and near organelles, including reservosomes (Fig. 5Ca-d). Parasites incubated with 200 μM CdTe QDs showed several alterations. Fig. 5E-G shows evidence of abnormal chromatin condensation, suggesting DNA damage and intense cytosolic vacuolisation. Endoplasmic reticulum profiles surrounding subcellular structures and myelin-like figures were indicative of autophagic processes (Fig. 5F). This concentration led to mitochondrial swelling (Fig. 5G) with loss of its matrix and, occasionally, total disruption of the organelle. We also observed QDs in membrane vesicles close to the flagellar region (Fig. 5H) and extensive changes in the plasma membrane near the flagellar pocket (Fig. 5I), suggesting the occurrence of membrane shedding. It is important to mention that no change was observed in the flagellum and sub-pellicular microtubule layers of these parasites (Fig. 5H, I). DISCUSSION QD toxicity has been extensively studied in various cellular prokaryote and eukaryote cellular models (Derfus et al. 2004, Pelley et al. 2009). The potential effects of CdTe in T. cruzi epimastigotes were evaluated in this work. This analysis involved parasite proliferation under different concentrations of CdTe QDs. High doses of QDs (20 and 200 μM) led to a decrease in T. cruzi growth patterns. However, lower concentrations presented similar proliferative rates to the control. Derfus et al. (2004) noted the toxicity of CdSe functionalized with MAA in primary hepatocytes. The authors showed that CdSe at low concentrations (0.25 mg/mL), which is equivalent to more than 20 μM CdTe used in this study, was not toxic to cells and caused no deleterious effects on hepatocellular function over seven days. The concentrations used here to label T. cruzi and intestinal epithelial cells of R. prolixus (2 and 10 μM of CdTe and CdSe) were lower than those previous employed (Derfus et al. 2004), encouraging further study into the interaction between parasites and vertebrate host cells. Lovri et al. (2005b) reported CdTe toxicity in murine microglia cells, demonstrating that high concentrations of QDs with green emission (10-100 μg/mL) induced cellular changes such as chromatin condensation, disorder of the nuclear membrane and a decrease in cellular metabolism. Our cell viability data showed that T. cruzi incubated with 200 μM CdTe do not die by necrosis within 72 h (Fig. 3). As previously described (Menna-Barreto et al. 2007), PI labelling was employed to evaluate the cell cycle of the parasite. PI is a well-known DNA marker; extensive studies relating the labelling intensity and different amounts of nucleic acids have been published. Intense labelling represented parasites with duplicated DNA; low labelling (half the intensity of the high-labelled samples) showed parasites in interphase and no labelling represented debris and/or G0 cells. In this work, the cell-cycle analysis showed that high concentrations (200 μM) of QDs decreased the percentage of duplicated-DNA parasites (14.5%) and increased the percentage of fragmented-DNA parasites (36.7%, as compared with control groups (36.6% and 11.9%, respectively) (Fig. 4). These results suggest that high concentrations of CdTe QDs lead to DNA changes in T. cruzi, compromising the proliferation of the parasite (Fig. 2). Some authors have observed alterations in DNA caused by nanoparticles, including CdTe aggregation in the nucleus (Weis 2003, Lovri et al. 2005b). It has already been shown that QDs capped with urea and acetate have affinity for the nuclei of mouse fibroblast cells (Bruchez et al. 1998). Nanoparticle size is also an essential factor in the analysis of toxicity because smaller CdTe QDs with green emission (2 nm) seem to be more toxic to the cells than those with red emission (6 nm) (Lovri et al. 2005b, Zhang et al. 2007). Our ultrastructural data showed that 2 μM QDs induced no alteration in T. cruzi morphology (Figs 3, 4). However, several important injuries were observed when parasites were incubated with 200 μM QDs. The extensive mitochondrial swelling observed was verified by Cho et al. (2007) who reported a similar effect when human breast cancer cells were incubated 10 mg/mL CdTe-Cys (about 100 μM CdTe). The authors also observed abnormal chromatin condensation (Cho et al. 2007). Morphological data, such as endoplasmic reticulum profiles surrounding the subcellular structure and myelin-like structures, led us to suggest that autophagy is occurring. As reviewed by Pelley et al. (2009), the toxicity of QDs is associated with their physicochemical properties. Due to the large diversity of nanoparticles (CdTe, CdSe, indium phosphide) and different capping techniques (MAA, zinc sulfide, MAS) employed, it is not possible to elucidate all possible toxicity mechanisms. The concentrations used in each model vary greatly. Several attempts have been made to reduce particle size by (i) selecting the capping of the nanoparticles, (ii) using minimal doses and (iii) modulating nanoparticle size (Bruchez et al. 1998, Hardman 2006, Cho et al. 2007, Tang et al. 2008). All of these factors are summarized in Table. Most studies on the mechanisms of QD toxicity were performed over short periods of time of up to 48 h (Pelley et al. 2009). In contrast, in the present work, several experiments were performed for longer periods of time. Parasites incubated with 20-200 μM CdTe QDs for seven days displayed changes in cell viability, cell cycle and ultrastructure after 72 h. In previous studies, our group demonstrated the presence of fluorescence in parasite-vector interactions followed for up to 11 days (Feder et al. 2009). Therefore, we showed that MAA-capped CdTe at concentrations up to 10 μM were able to mark T. cruzi and insect vector cells successfully. Taken together, these results demonstrate that a low concentration of CdTe QDs (2 μM) is optimal for use in bioimaging, whereas high concentrations (200 μM CdTe) could be toxic to cells and cause morphological changes. Corroborating these results, transmission electron microscopy results demonstrated the presence of QDs in cytoplasmic vesicles, dispersed in the cytoplasm and near reservosomes. These results suggest that nanoparticles are being endocytosed via an unknown mechanism. We did not observe the presence of QDs within the reservosomes, showing that they are probably not stored in this organelle as suggested by confocal microscopy results. In addition, we verified the presence of QDs around lipid droplets and dispersed in the cytoplasm (data not shown), indicating that QDs not conjugated to specific molecules may be located in different compartments. Some studies have shown that the size of the nanoparticle influences its subcellular distribution. Smaller QDs tend to accumulate in the nucleus whereas larger QDs remain in the cell cytoplasm (Lovri et al. 2005b, Hardman 2006). Studies on the ejection of QDs by cells are rare and are usually conducted in mammalian models (Chen et al. 2008, Lin et al. 2008); studies on the elimination of QDs by microorganisms are very scarce. Our results indicate a possible T. cruzi pathway for the elimination of QDs; their presence in vesicles and the plasma membrane shedding data suggest that QDs could be externalized by these mechanisms. Vesicles labelled with QDs were also observed surrounded by profiles of the endoplasmic reticulum, indicating an autophagic process that may be an attempt to eliminate the QDs. In the present study we demonstrated that T. cruzi labelled with low concentrations of CdTe QDs did not affect (i) parasite integrity up to seven days, (ii) parasite cell division or (iii) parasite motility. This fact was reinforced by the low level of QD cytotoxicity in the parasites. In summary, our results show that T. cruzi labelled with 2 μM of QDs do not experience toxic effects and that these QDs are suitable for in vivo cellular studies. REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11026f1.jpg] [oc11026t1.jpg] [oc11026f3.jpg] [oc11026f2.jpg] [oc11026f5.jpg] [oc11026f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}