 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
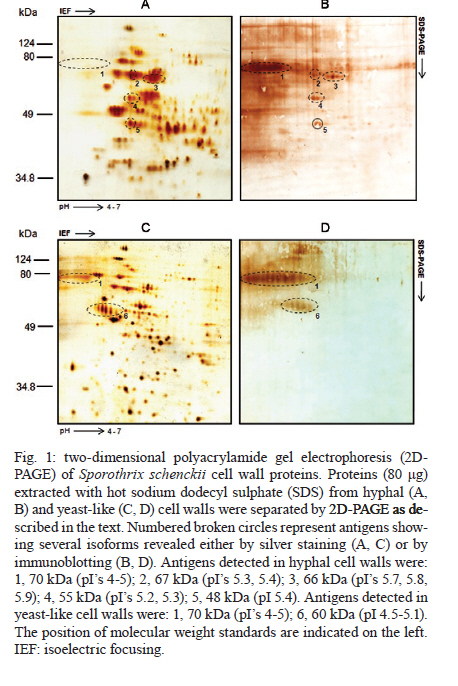
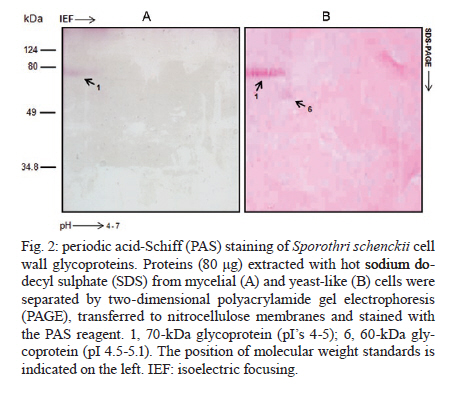
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 2, 2011, pp. 248-250 SHORT COMMUNICATIONS 2D-immunoblotting analysis of Sporothrix schenckii cell wall Estela Ruiz-BacaI, +; Héctor M Mora-MontesII; Everardo López-RomeroII; Conchita TorielloIII; Virgilio Mojica-MarínI; Norma Urtiz-EstradaI IFacultad
de Ciencias Químicas, Universidad Juárez del Estado de Durango,
Durango, México + Corresponding author: erb750@hotmail.com Received 7 August
2010 Code Number: oc11040 ABSTRACT We utilized two-dimensional gel electrophoresis and immunoblotting (2D-immunoblotting) with anti-Sporothrix schenckii antibodies to identify antigenic proteins in cell wall preparations obtained from the mycelial and yeast-like morphologies of the fungus. Results showed that a 70-kDa glycoprotein (Gp70) was the major antigen detected in the cell wall of both morphologies and that a 60-kDa glycoprotein was present only in yeast-like cells. In addition to the Gp70, the wall from filament cells showed four proteins with molecular weights of 48, 55, 66 and 67 kDa, some of which exhibited several isoforms. To our knowledge, this is the first 2D-immunoblotting analysis of the S. schenckii cell wall. Key words: sporotrichosis - Sporothrix schenckii - cell wall - glycoproteins - 2D-immunoblotting analysis The dimorphic fungus Sporothrix schenckii is the etiological agent of sporotrichosis, an endemic subcutaneous mycosis in Latin America, whose prevalence has significantly increased during the last few years mainly in immunocompromised patients (Callens et al. 2006, Lopes-Bezerra et al. 2006). The S. schenckii cell wall is composed of alkali-soluble and insoluble glucans, which are found in both morphological phases of the fungus (Previato et al. 1979, Lopes-Bezerra et al. 2006). Despite significant progress in the knowledge of structural polysaccharides, little is known regarding the identity and characteristics of cell wall proteins. Thus far, a peptide-rhamnomannan and peptide-rhamnogalactan have been isolated and characterized from wall preparations of yeast-like cells (Lloyd & Bitoon 1971, Nakamura 1976, Lopes-Bezerra et al. 2006). In addition, there are some reports dealing with the role of a cell wall glycoprotein of 70 kDa in adhesion of S. schenckii to host tissues and extracellular matrix components as well as fungal pathogenesis (Nascimento et al. 2008, Ruiz-Baca et al. 2009, Teixeira et al. 2009). 2D-immunoblotting analysis has become one of the most frequently used tools to search antigens present in fungal pathogens such as Candida albicans, Cryptococcus neoformans and Aspergillus fumigatus (Pitarch et al. 2002, 2009, Gautam et al. 2007, Young et al. 2009). This approach was used here to further advance in the identification of antigenic proteins present in the cell wall of both morphologies of S. schenckii. Antibodies raised against the whole fungal cell allowed the identification of morphology-specific cell wall glycoproteins as well as glycoproteins present in both fungal phases. Organism and culture conditions - S. schenckii ATCC 58251 was used in this study. For mycelia propagation, 2-l Erlenmeyer flasks containing 600 mL of YPG medium (0.3% yeast extract, 1% peptone and 2% glucose, pH 4.5) were inoculated with 1 x 106 conidia mL-1 and incubated for 24 h at 28ºC with shaking (120 rpm). To obtain the yeast-like form, the pH of the medium was adjusted to 7.2 and the flasks were shaken for six days at 37ºC. Hyphae were collected by filtration in a Büchner filter and yeast-like cells were harvested by centrifugation at 7,000 g for 10 min. In both cases, cells were washed twice with lysis buffer (50 mM Tris-HCl buffer, pH 7.5, supplemented with 1 mM phenylmethylsulfonyl fluoride). Extraction of cell wall proteins - Mycelia and yeast-like cells were resuspended in 10 mL of lysis buffer and disrupted in an MSK cell homogenizer (Braun Melsungen, Germany) as described previously (Mora-Montes et al. 2010). To isolate cell walls, the cell homogenate was centrifuged for 15 min at 1,300 g and 4ºC and the pellet was saved and washed five times with lysis buffer to eliminate any intracellular component associated with the walls during the cell disruption process. Cell wall proteins were extracted with hot 2% sodium dodecyl sulfate (SDS) as previously described (Ruiz-Baca et al. 2009). Proteins were precipitated with 70% (v/v) ethanol at -20ºC for 5 h and kept at -70ºC until use. Protein concentration was determined by the Bradford method (1976). Two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) - Samples containing 80 μg of protein were cleaned with the ReadyPrepTM 2D-Cleanup kit (Bio-Rad) following the manufacturer's instructions, solubilized in 125 μL of rehydration buffer (7 M urea, 2 M thiourea, 4% CHAPS, 20 mM DTT, 0.5% ampholytes and 0.002% bromophenol blue) and applied onto Immobiline pH 4-7 non-linear DryStrips (7 cm long, Bio-Rad). Strips were rehydrated for 16 h before isoelectric focusing (IEF). IEF was performed with a Protean IEF Cell (Bio-Rad) at 20ºC using the following conditions: 250 V for 15 min, 250-4,000 V for 2 h and 4,000 V until complete 10,000 V/h. After IEF, strips were sequentially equilibrated for 15 min each in the equilibrium solution (50 mM Tris-HCl, pH 8.8, 6 M urea, 30% glycerol, 2% SDS, 0.5% DTT and 2% iodoacetamide) with constant agitation. For the second dimension, IEF strips were placed on top of a 10% vertical SDS-PAGE gel and covered with 0.5% agarose. Proteins were separated for 1 h at 90 V in a Mini-PROTEAN 3 Cell unit (Bio-Rad) and either silver stained (Merril 1990) or transferred to nitrocellulose membranes. Images were captured using an Image Scanner (Bio-Rad) and analyzed with the PDQuestTM 2-D analysis software (Bio-Rad). Assays were repeated at least thrice using independent cell wall preparations. Antibodies - Anti-S. schenckii antibodies were raised in a New Zealand male rabbit following the immunization scheme described by Toriello and Mariat (1974). Briefly, 1 mL aliquots containing 1 x 108 formaldehyde-killed yeast-like cells were inoculated into the marginal ear vein of the animal three times per week for two weeks and one week after the final injection the rabbit was bled. The blood serum was saved and stored at -70ºC in aliquots of 2 mL containing 0.01% (w/v) sodium azide. Immunoblotting assays - Immunodetection was carried out as previously described (Mora-Montes et al. 2008), except that a primary polyclonal anti-S. schenckii antibody diluted 1:500 in phosphate buffered saline-Tween 20 (0.05%) was used. Glycoprotein staining - Proteins transferred to nitrocellulose membranes after 2D-PAGE were stained with periodic acid-Schiff (PAS) reagent as described previously (Ruiz-Baca et al. 2009). Extraction of cell walls with hot SDS rendered significantly more protein from yeast-like cells (2.1 ± 0.02 μg per mg of cell wall dry weight) than from mycelia (1.2 ± 0.03 μg per mg of cell wall dry weight). Silver staining after 2D-PAGE revealed a higher number of proteins extracted from hyphal walls (Fig. 1A) than from yeast walls (Fig. 1C) (130 vs. 115 protein spots, respectively). A 70-kDa protein showing several isoforms in a pI range from 4-5 was immunodetected in extracts of walls from both morphologies (Fig. 1B, D). In addition, hyphal walls presented proteins with molecular masses of 48, 55, 66 and 67 kDa (Fig. 1B). Moreover, a 60-kDa protein with several isoforms in a pI range from 5-6 was immunorecognized only in yeast walls (Fig. 1D). To determine whether some of the immunodetected proteins were glycosylated, the 2D-PAGE gels were stained with PAS reagent. Results indicated that both the 60 and 70-kDa proteins present in yeast walls were glycoproteins and that the former was present only in extracts of hyphal walls (Fig. 2). It should be noted that the presence of other glycoproteins in the morphological phases of the fungus cannot be ruled out as the methodology used here detects only proteins with a rather high degree of glycosylation. The antigenic composition of both morphologies of S. schenckii is poorly understood and there are only a few described antigens consisting mainly of glycoproteins with a high degree of glycosylation, containing mannose, rhamnose and galactose. The sugar composition of glycoproteins varies with the culture age and is closely related to the growth morphology of the organism (Takata & Ishizaki 1983). Scott and Muchmore (1989) demonstrated that yeast-like cells of S. schenckii contain two proteins of 40 and 70-kDa as the major antigens recognized by sera obtained from sporotrichosis patients; also, it was demonstrated that sera from S. schenckii-infected mice reacted mainly with a 70-kDa antigen (Nascimento & Almeida 2005). In addition, we have purified and characterized a 70-kDa glycoprotein (Gp70) from the cell wall of S. schenckii yeast-like cells that exhibited adhesion properties (Ruiz-Baca et al. 2009). Blotting with a polyclonal anti-Gp70 serum revealed a single spot with a pI of 4.1, which suggested that Gp70 is a S. schenckii-specific protein as it was not immunodetected in other pathogenic and non-pathogenic fungi (Ruiz-Baca et al. 2009). Similar studies have not been carried out in the filamentous phase. Almeida-Paes et al. (2007) identified five exoantigens of 42, 51, 63, 70 and 90 kDa in Sabouraud dextrose cultures of mycelial cells of S. schenckii 23508, with the 51 and 63 kDa antigens being the most prominent. Optimum expression of these components occurred in the stationary phase of growth. Here, five antigens of 48, 55, 66, 67 and 70 kDa were immunodetected in the mycelial phase of S. schenckii ATCC 58251, whereas only two antigens of 60 and 70 kDa were identified in yeast cells. Because mycelial cell antigens have not been purified or characterized, more studies will be necessary to verify if these are related to those previously reported and to determine their presence in other fungal strains. The observed differences between yeast and mycelial cells in terms of the proteins detected in this study may result from the physicochemical nature of the proteins obtained from each form. The amount of acidic and basic proteins in the two morphologies may be different, with the mycelial phase containing more proteins resolved within the pH 4-7 range (Fig. 1A). In summary, the Gp70 was the major antigenic protein present in the walls of both fungal morphologies and this may correspond to the previously characterized Gp70. As the S. schenckii genome has not been yet sequenced, 2D-immunoblotting studies focused on surface antigens can help to understand many of the molecular aspects that govern the biology of the fungus and sporotrichosis. ACKNOWLEDGEMENTS To Bertha C Calderón Barraza, for technical assistance. REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11040f2.jpg] [oc11040f1.jpg] |
| |||||||||

{kind=link}
{kind=link}