 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 2, 2011, pp. 251-253 SHORT COMMUNICATIONS In vitro activity of amphotericin B cochleates against Leishmania chagasi Aretha Molina Sesana; Renata Monti-Rocha; Solange Alves Vinhas; Carlos Gustavo Morais; Reynaldo Dietze; Elenice Moreira Lemos+ Núcleo de Doenças Infecciosas, Universidade Federal do Espírito Santo, Vitória, ES, Brasil + Corresponding author: lemosem@ndi.ufes.br Received 30 August
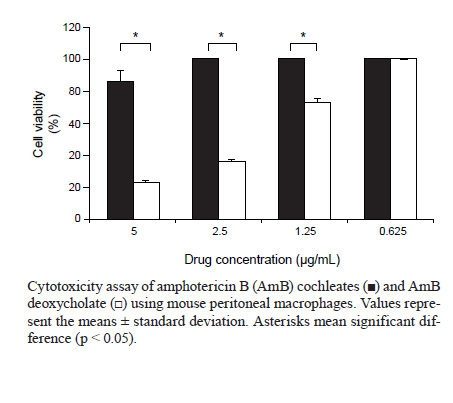
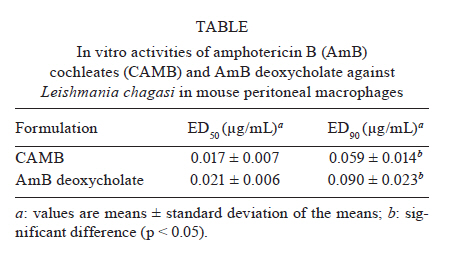
2010 Code Numbner: oc11041 ABSTRACT Cochleate delivery vehicles are a novel lipid-based system with potential for delivery of amphotericin B (AmB). In this study, the efficacy of cochleates was evaluated by examining the in vitro activity of AmB cochleates (CAMB) against Leishmania chagasi in a macrophage model of infection. We demonstrate that CAMB is nontoxic to macrophages at concentrations as high as 2.5 μg/mL, whereas the conventional formulation, AmB deoxycholate, showed high toxicity at this concentration. The in vitro activity of CAMB against L. chagasi was found to be similar to that of the reference drug AmB deoxycholate, with ED50s of 0.017 μg/mL and 0.021 μg/mL, respectively. Considering that L. chagasi affects organs amenable to cochleate-mediated delivery of AmB, we hypothesize that CAMB will be an effective lipid system for the treatment of visceral leishmaniasis. Key words: amphotericin B cochleates - Leishmania chagasi - in vitro activity Despite its potent activity against leishmaniasis, the toxicity of the conventional formulation of amphotericin B (AmB) complexed with deoxycholate micelles limits its use in the treatment of visceral leishmaniasis (VL). To improve the therapeutic index of AmB and reduce its toxicity, new lipid-based formulations have been developed (Robinson & Nahata 1999). These drug delivery systems, such as liposomal formulations, lipid complexes, lipid emulsions and colloidal dispersions, have been introduced into clinical practice. However, despite their proven success against leishmaniasis (Dietze et al. 1993, Davidson et al. 1996, Sundar et al. 1997) their high cost impedes their frequent use. Cochleate delivery vehicles are a novel lipid-based system that has potential for AmB delivery. As described by Zarif et al. (2000), cochleates have a unique multilayered structure consisting of a large, continuous, solid lipid bi-layer sheet rolled into a spiral, with no internal aqueous space. Cochleates are stable phospholipid-calcium precipitates comprised mainly of phosphatidylserine, with a mean diameter of 407 ± 233.8 nm and molar ratio of 10:1 lipid/AmB when in complex with the drug. Because the entire cochleate structure is a series of solid layers, components within the interior of the cochleate structure are protected from exposure to harsh environmental conditions or enzymes. Given this feature, cochleates should be an ideal system for oral delivery of AmB. Beside their usefulness as a system to deliver protein, peptide and DNA for vaccine and gene therapy applications (Mannino & Gould-Fogerite 1997, Zarif & Mannino 2000), cochleates have also been used as a delivery system for a variety of drugs, including AmB (Santangelo et al. 2000, Zarif et al. 2000, Delmas et al. 2002, Syed et al. 2008, Livne et al. 2010). Additionally, a comprehensive patent specifying that Leishmania major-infected mice were orally treated with AmB cochleates (CAMB) was recently published (Mannino 2010). In the present study, the efficacy of cochleates as a delivery vehicle for AmB was evaluated by examining the in vitro activity of CAMB against Leishmania chagasi in a macrophage model of infection. Its activity and toxicity were also compared to a commercially available AmB preparation. CAMBs were developed as described by Zarif et al. (2000) and provided by BioDelivery Sciences, USA. Amphotericin B deoxycholate (Fungizone™, Bristol Myers Squibb, France) were purchased commercially in 50 mg vials and were reconstituted according to the manufacturer's instructions. For in vitro experiments, the drugs were prepared by diluting the stock solutions in RPMI 1640 medium containing 10% of foetal calf serum according to the concentrations required on the day of experiment. To assess the cytotoxicity of CAMB, peritoneal macrophages were seeded at 8 x 104 cells/well in 96-well microplates and incubated overnight with a supply of 5% CO2 at 37ºC. CAMB and AmB deoxycholate at different concentrations (5-0.625 μg/mL) were dispensed in triplicate, with three wells without drug acting as the control. The plates were incubated for 72 h and, to assess cell viability, an 3-(4,5-dimethylthiazole-2-yl)-2-5-diphenyl tetrazolium bromide (MTT) assay was performed. Briefly, 100 μL MTT (0.5 mg/mL) was added to each well and incubated for 4 h. The plate was spun down at 1,000 g for 10 min at 4ºC and the supernatant was aspirated. Yellow tetrazolium salt that had reduced to blue formazan within cells was then dissolved in 100 μL of dimethyl sulfoxide and the optical density at 570 nm was determined. The results were expressed as percentage of viable cells compared with untreated control wells. The in vitro efficacy of CAMB against L. chagasi was evaluated by amastigote-macrophage assay. Peritoneal macrophages of Swiss mice were plated in Labtek 16-well tissue culture slides (Nunc, NY, USA) and were allowed to adhere for 24 h in an incubator with a supply of 5% CO2 at 37ºC. Adherent macrophages were infected with late-log phase promastigotes of L. chagasi (MHOM/BR/74/PP75) at a ratio of 7:1 promastigotes: macrophage. After 24 h of incubation, free promasti-gotes were removed and the culture was exposed to drugs over a dose range of 0.078-0.002 μg/mL, in triplicate at each concentration. After 72 h of additional incubation, the slides were fixed and stained with Panótico solutions (Laborclin, Paraná, Brazil). At least 100 cells were counted to determine the percentage of infected macrophages. The percent killing was calculated by sigmoidal regression analysis (Prisma 5.03) and the mean ED50s and ED90s were determined in three independent assays. An unpaired t-test was applied to determine the significance of CAMB ED50 and ED90 over AmB deoxycholate in the inhibition of infection in macrophages. In this study, the toxicity of CAMB on mouse peritoneal macrophages was analyzed and compared to that found for AmB deoxycholate. The percentage of live cells in cultures treated with CAMB at 5 μg/mL was 86%, while treatment with AmB deoxycholate at the same concentration resulted in 100% killing. At concentrations of 2.5 μg/mL CAMB had no toxicity for macrophages, while the same concentration of AmB deoxycholate showed high toxicity, with only 36% of cells surviving. AmB deoxycholate was non-toxic at a concentration of 0.625 μg/mL, demonstrating its high toxicity (Figure). Cochleates lacking drug at 5 μg/mL were non-toxic to cells and there was no difference in cell viability between treated and untreated cells. The in vitro activity of CAMB against intracellular L. chagasi amastigotes was found to be similar to that of the reference drug AmB deoxycholate, with an ED50 of 0.017 μg/mL for CAMB and 0.021 μg/mL for AmB deoxycholate. However, CAMB showed higher efficacy compared to the control drug when ED90s was evaluated (Table). Empty cochleates did not show any anti-leishmanial activity. Different studies suggest that drugs incorporated into liposome or lipid formulations are selectively taken up by the phagocytic cells and are concentrated in the liver, spleen and lung (Hiemenz & Walsh 1996). Moreover, it has been demonstrated that AmB liposomal products have significantly less nephrotoxicity than conventional amphotericin B, even at much higher doses (Walsh et al. 1998). Encapsulation of AmB into cochleates results in stable, nontoxic and highly efficacious AmB lipid particles, facilitating systemic delivery of AmB. In our study, CAMB treatment was found to be nontoxic to macrophages even at concentrations as high as 2.5 μg/mL, whereas AmB desoxycholate showed high toxicity at this same concentration, confirming the lower toxicity of this system compared to the commercial formulation. In the CAMB formulation, cells do not interact with free AmB, making CAMB less toxic than desoxycholate AMB. Intact cochleates can induce a perturbation and reordering of the cell membrane, resulting in fusion between the outer layer of the cochleate and the cell membrane. This fusion promotes the delivery of a small amount of the encochleated drug to the cytoplasm of the target cell. Cochleates can also be endocytosed into the cytoplasm of the cell. Once within the low calcium environment of the cytoplasm, the structure of the cochleates alters and the drug is released (Mannino 2010). In addition to its efficacy in the treatment of experimental candidiasis and aspergillosis (Santangelo et al. 2000, Zarif et al. 2000, Delmas et al. 2002), our results showed that CAMB has a substantial inhibitory action on the growth of L. chagasi amastigotes. Because of the hydrophobicity of AmB molecules, they integrate particularly well with the cochleate struture, providing the additional benefit of protecting AmB from exposure to harsh environmental conditions or enzymes and facilitating oral administration. Pharmacokinetic studies confirmed that following multiple oral administrations of CAMB, AmB is distributed in target tissues, with the highest concentration of AmB being in the kidneys, followed by the lungs, liver and spleen (Delmas et al. 2002). The ability of cochleates to deliver systemic AmB following multiple oral doses highlights the potential of CAMB formulations to treat and prevent invasive infections. In summary, our results demonstrate that CAMB represents a potential lipid-based vehicle for AmB delivery, with high in vitro activity against L. chagasi and lower toxicity when compared to the conventional formulation of AmB deoxycholate. Considering that L. chagasi infection affects organs targeted by cochleate-mediated delivery of AmB, we can hypothesize that CAMB could be used as a lipid system to treat VL. To investigate this hypothesis, we will further evaluate the potential of CAMB to treat experimental VL using the oral route of administration. ACKNOWLEDGEMENTS To Dr Raphael Mannino, from BioDelivery Science, for providing the amphotericin B cochleates. REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11041t1.jpg] [oc11041f1.jpg] |
| |||||||||

{kind=link}
{kind=link}