 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 3, 2011, pp. 408-414 ARTICLES The effect of combined polymorphisms in chemokines and chemokine receptors on the clinical course of HIV-1 infection in a Brazilian population Valdimara Corrêa VieiraI; Maria Fernanda Martínez BarralI; Raul Andrés Mendoza-SassiI; Jussara Maria SilveiraI; Marcelo Alves SoaresII, III, +; Ana Maria Barral de MartínezI IFaculdade
de Medicina, Universidade Federal do Rio Grande, Rio Grande, RS, Brasil Financial support: CNPq (471207/2008-0), FAPERJ (E-26/220.336/2007) Received 25 October
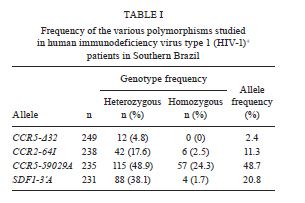
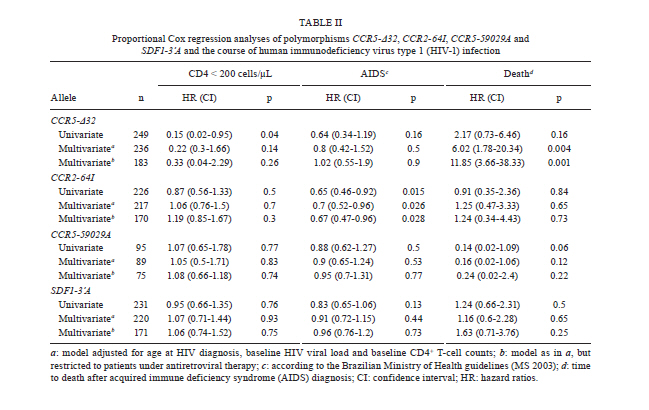
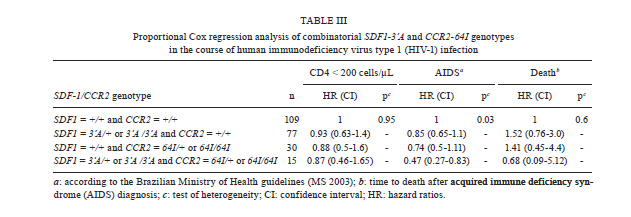
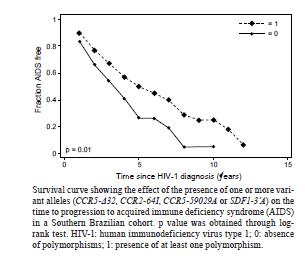
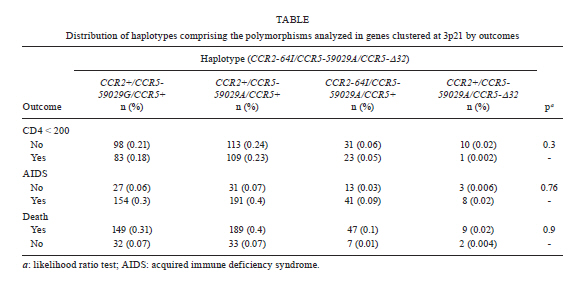
2010 Code Number: oc11071 Abstract Polymorphisms in genes that encode chemokines or their receptors can modulate susceptibility to human immunodeficiency virus (HIV) infection and disease progression. The objective of this study was to assess the frequency of polymorphisms CCR5-Δ32, CCR2-64I, CCR5-59029A and SDF1-3'A and their role in the course of HIV infection in a Southern Brazilian population. Clinical data were obtained from 249 patients for an average period of 6.4 years and genotypes were determined by standard polymerase chain reaction (PCR) and PCR-restriction fragment length polymorphism. Survival analyses were conducted for three outcomes: CD4+ T-cell counts below 200 cells/µL, acquired immune deficiency syndrome (AIDS) or death. The frequency of the polymorphisms CCR5-Δ32, CCR2-64I, CCR5-59029A and SDF1-3'A were 0.024, 0.113, 0.487 and 0.207, respectively. CCR5-Δ32 was associated with a reduction in the risk for CD4+ T-cell depletion and with an increased risk for death after AIDS diagnosis. CCR2-64I was associated with a reduction in the risk for developing AIDS. SDF1-3'A was also associated with decreased risk for AIDS, but its effect was only evident when CCR2-64I was present as well. These results highlight the possibility of using these markers as indicators for the prognosis of disease progression and provide evidence for the importance of analysing the effects of gene polymorphisms in a combined fashion. Key words: CCR5-Δ32 - CCR2-64I - CCR5-59029A - SDF1-3'A- HIV - disease progression A significant portion of the variable clinical course profiles observed in the natural history of human immunodeficiency virus type 1 (HIV-1) infection is attributed to host genetic factors (Dean et al. 1996, Borghans et al. 2007, van Manen et al. 2008). Among these genetic factors are mutations found in genes encoding chemokines or their receptors (Dean et al. 1996, Quillent et al. 1998), which are key molecules in the HIV life cycle (Cocchi et al. 1995, Doranz et al. 1996, Dragic et al. 1996, Feng et al. 1996). One of the most notable effects on the clinical course of HIV infection is conferred by an internal 32 bp deletion (∆32) in the CCR5 chemokine receptor gene (CCR5-∆32). This mutation leads to the synthesis of a truncated protein that fails to reach the cell surface (Liu et al. 1996). Homozygous individuals (∆32/∆32) are highly resistant to R5-tropic HIV-1 viruses (Dean et al. 1996, Samson et al. 1996). Heterozygous subjects (+/∆32) are susceptible to infection but exhibit slower rates of progression to acquired immune deficiency syndrome (AIDS) when compared to wild-type homozygous counterparts (Dean et al. 1996, Liu et al. 1996, Samson et al. 1996). The frequency of the CCR5-∆32 allele varies in different ethnicities, occurring at a rate of 10-15% in Caucasians, 7% in Hispanics and 6% in African Americans, while it is virtually absent in native Africans and Asians (Samson et al. 1996, Williamson et al. 2000, Wang et al. 2003). Polymorphisms in the promoter region of the CCR5 gene, including a G/A polymorphism at position -59029, have also been correlated with alterations in the clinical course of HIV infection (Martin et al. 1998, McDermott et al. 1998, An et al. 2000, Kaur et al. 2007). McDermott et al. (1998) showed that individuals homozygous for the -59029A variant progress faster to AIDS when compared to subjects homozygous for the -59029G allele. The effect was stronger in the absence of the protective alleles CCR5-∆32 and a non-synonymous amino acid polymorphism found at codon 64 of the CCR2 receptor gene (CCR2-64I). In the same study, it was also shown that the promoter region for the -59029G allele displays 45% reduced activity compared to that of the -59029A variant in vitro. In a study conducted with both Caucasian and African-American patients, the -59029A allele was associated with a rapid progression to AIDS in both ethnicities. This phenotype was recessive in the former group and dominant in the latter (An et al. 2000). Both alleles are common, with the occurrence rate of -59029A varying from 43-68%, depending on ethnicity (McDermott et al. 1998). The effects of specific CCR5 haplotypes, consisting of polymorphisms in the CCR5/CCR2 promoter and coding regions, have also been reported. Particularly, the CCR5 HHE/HHE and HHC/HHG*2 genotypes have been associated with, respectively, detrimental and beneficial effects in the course of HIV infection (Gonzales et al. 1999, Mangano et al. 2001, Nguyen et al. 2004, Casado et al. 2010, Catano et al. 2011). Polymorphisms in CCR5 as well as in other chemokine and chemokine receptor genes have also been associated with altered susceptibility to other infectious diseases, such as Chagas disease and hepatitis C (Hellier et al. 2003, Cunha-Neto et al. 2009). A point mutation described in the gene encoding the CCR2 chemokine receptor also plays a role in HIV-1 pathogenesis. The polymorphism CCR2-64I represents a conservative change in the first transmembrane domain of the protein (Smith et al. 1997, Kostrikis et al. 1998), which does not alter its properties as a chemokine or HIV-1 receptor (Lee et al. 1998). Several studies have suggested a better prognosis for AIDS patients carrying the CCR2-64I variant (Smith et al. 1997, Kostrikis et al. 1998, Mulherin et al. 2003, Passam et al. 2005), but others have found contradictory results (Michael et al. 1997, Schinkel et al. 1999). Mummidi et al. (1998) showed a protective effect of this allele in African-Americans but not in Caucasians. Unlike the CCR5-∆32 mutation, CCR2-64I was present in all ethnicities tested. CCR2-64I was observed at a frequency of 10% in Caucasians, 15% in African-Americans, 17% in Hispanics and 25% in Asians (Smith et al. 1997, Williamson et al. 2000, Wang et al. 2003). The polymorphism SDF1-3'A is characterised by a G-to-A transition in the 3'-UTR of the gene and was initially correlated with protection from HIV disease progression (Winkler et al. 1998). However, other studies did not find such a correlation (Ioannidis et al. 2001, Watanabe et al. 2003, Reiche et al. 2006), while a third group of studies found a correlation with accelerated disease progression (Mummidi et al. 1998, van Rij et al. 1998, Daar et al. 2005). SDF1-3'A distribution varies significantly between different populations, with frequencies from 2-9% in Africans and up to 54-71% in Oceania (Su et al. 1999). CCR5-∆32, CCR2-64I, CCR5-59029A and SDF1-3'A all exhibit variable frequencies in distinct populations (Smith et al. 1997, McDermott et al. 1998, Su et al. 1999, Gonzalez et al. 2001) and, possibly, their phenotypes depend on the ethnicity analysed (Mummidi et al. 1998, Gonzalez et al. 1999, 2001, An et al. 2000). The Brazilian population presents a complex genetic background, characterised by a high degree of miscegenation (Callegari-Jacques et al. 2003, Pena 2005). In major Brazilian cities, CCR5-∆32 was found at frequencies of between 2-7% (Munerato et al. 2003, Vargas et al. 2006, Reiche et al. 2008, Rigato et al. 2008), CCR5-59029A between 39-44% (Rigato et al. 2008), CCR2-64I at approximately 10% (Munerato et al. 2003, Rigato et al. 2008) and SDF1-3'A between 18-22% (Reiche et al. 2006, Rigato et al. 2008). Despite the moderate rates of HIV infection reported in Brazil, little is known about the influence of genetic polymorphisms on HIV disease progression in this population. The aim of this study was to determine the frequency of the alleles CCR5-∆32, CCR2-64I, CCR5-59029A and SDF1-3'A and their combined influence on HIV disease progression in a cohort of HIV-infected patients from Southern Brazil. Patients, Materials and Methods Subjects and data collection - This was a retrospective longitudinal study. Two hundred and forty-nine HIV-1+ patients followed at the University Hospital of Universidade Federal do Rio Grande (HU-FURG) were recruited. Patients were followed for an average duration of 6.44 years. Clinical and laboratory data were obtained through medical chart review for the complete period between the diagnosis of HIV infection and the last follow-up visit until recruitment to the study. Baseline variables assessed were HIV plasma viral load, CD4+ T-cell counts and clinical status. This study was approved by the Committee on Ethics in Institutional Research (CEPAS-FURG 23.116.002593/2007- 04). Molecular analyses - Genomic DNA was extracted from patient whole blood samples using the GFX Genomic Blood PurificationTM kit (Amersham Biosciences, Piscataway, NJ, USA). Genotyping of the CCR5 coding region (rs333) was performed using polymerase chain reaction (PCR), while genotyping of CCR2 (rs1799864), the CCR5 promoter (rs1799987) and SDF1 (rs1801157) were performed using PCR followed by restriction fragment length polymorphism (PCR-RFLP). PCR reactions were conducted in a final volume of 25 µL consisting of 5 pmol of each primer, 0.3 mM dNTP, 2.5 mM MgCl2, 0.75 U of Taq polymerase (Invitrogen Corporation, San Diego, CA), 20 mM Tris-HCl (pH 8.4), 50 mM KCl and approximately 0.5 ug of genomic DNA. For the CCR5 coding region, the primers 5'CAAAAAGAAGGTCTTCATTACACC3' (forward) and 5'CCTGTGCCTCTTCTCATTTCG3' (reverse) were used as previously described (Huang et al. 1996). For the wild type allele, the amplified product was 189 bp long, while for the CCR5-∆32 allele, it was 157 bp long. PCR fragments were electrophoresed in a 2% agarose gel. For CCR2 PCR amplification, the primers 5'GGATTGAACAAGGACGCATTTCCCC3' (forward) and 5'TTGCACATTGCATTCCCAAAGACCC3' (reverse) were used as described previously (Magierowska et al. 1999). The 380 bp products were subjected to RFLP with the restriction enzyme BseGI, generating two fragments (215 and 165 bp) only when the mutation corresponding to CCR2-64I was present. The wild type allele remained uncut (Suresh et al. 2006). For the CCR5 promoter region, primers 5'AAAATCCCCACTAAGATCCTG3' and 5'ATTCATCTAGTCAAAAGCCCAC3' were used (An et al. 2000). The final product of 394 bp was digested with Bsp 1286 I, generating two (329 and 65 bp) or three (202, 127 and 65 bp) fragments for the alleles CCR5-59029A or CCR5-59029G, respectively. Finally, for the amplification of SDF-1 gene, the primers 5'-CAGTCAACCTGGGCAAAGCC-3' and 5'-CCTGAGAGTCCTTTTGCGGG-3' were used (Winkler et al. 1998). The 293 bp PCR product was subjected to RFLP with MspI. The digestion of the common SDF-1 allele produced two DNA fragments (100 and 193 bp), while the SDF1-3'A remained intact (Winkler et al. 1998). Statistical analyses - The frequencies of the allele mutations were calculated and the Hardy-Weinberg equilibrium adequacy was tested with X2. To evaluate the influence of the polymorphisms on the course of infection, Cox proportional analyses [hazard ratios (HR) and their respective 95% confidence intervals] were conducted for three endpoints: (i) decline of CD4+ T-cell counts to < 200 cells/µL, (ii) progression to AIDS according the Brazilian national guidelines (presence of at least one opportunistic disease and/or CD4+ T-cell counts < 350 cells/µL) and (iii) time to death after AIDS diagnosis. The single effect of each polymorphism was evaluated with univariate Cox regression. Because baseline CD4+ T-cell counts, baseline HIV viral load and age at HIV diagnosis could potentially act as confounding factors, multivariate analyses adjusted for these variables were performed for each polymorphism using Cox regression. A further multivariate analysis was conducted with the same model but included only patients undergoing antiretroviral therapy. The criterion for remaining in the model was a p value < 0.05. In view of the small number of observed individuals homozygous for the alleles CCR2-64I and SDF1-3'A, their effect was verified by comparing heterozygous and rare homozygous with the respective common homozygous allele for each locus (CCR2-64I/+ and CCR2-64I/64I x CCR2-+/+; SDF1-3'/+ and SDF1-3'A/3'A x SDF1-+/+). For CCR5-Δ32, heterozygous individuals were compared with wild-type homozygous patients (CCR5-Δ32/+ x CCR5-+/+) because no CCR5-Δ32/Δ32 patients were observed in the study. For CCR5-59029A, homozygotes for each allele (CCR5-59029A/A and CCR5-59029G/G) were compared with each other. Considering that the alleles CCR2-64I and CCR5-Δ32 constitute confounding factors in measuring the effect of CCR5-59029A (McDermott et al. 1998), subjects harbouring any of those alleles were excluded from the analyses for this polymorphism. Finally, analyses of subjects with different combinations of polymorphisms were performed. A survival curve was constructed and the level of significance was determined through a log-rank test. Data were analysed in Stata v. 9.0. Differences were considered to be significant when p < 0.05 in two-sided tests. Results Subject characteristics - Among the 249 patients included in the study, 163 (65.5%) were female. At the first visit after diagnosis, the mean age was 31.2 years [confidence interval (CI) 14.1-65.8], the mean CD4+ T-cell counts were 363 cells/µL [n = 248; standard deviation (SD) 277] and the average HIV viral load was 3.94 log10 copies/mL (n = 236; SD 2.48). At the beginning of follow-up, 117 (47%) patients were asymptomatic, 39 (15.7%) remained asymptomatic until the end of the study and 43 (17.3%) patients died. The average time from diagnosis to progression to AIDS was 1.76 years (SD 2.69) and to death was 5.88 years (SD 3.07). During the follow-up, 193 patients (83.9%) started antiretroviral therapy. Polymorphism frequencies and their influence on HIV disease progression - The frequency of the polymorphisms studied is reported in Table I. No CCR5-Δ32 homozygous subjects were observed. The genotypic frequencies observed were found to be in Hardy-Weinberg equilibrium (not shown). The results of Cox proportional analyses are shown in Table II. An HR > 1 or < 1 indicates a greater or lower risk of progression to the outcome analysed, respectively. In the univariate analysis, the single effect of the four polymorphisms was investigated. For the CCR5 coding region, the univariate analysis showed that individuals heterozygous for the allele (CCR5-∆32/+) presented an 85% decreased risk of progression to CD4+ counts below 200 cells/µL (HR = 0.15, p = 0.04) (Table II). However, in the multivariate analysis, this protective effect was not significant (Table II). Considering death as endpoint, CCR5-∆32/+ individuals had a significantly increased risk in the multivariate analysis (HR = 6.02, p = 0.004) (Table II). To assess the effect of the CCR2-64I allele in the absence of CCR5-∆32, we excluded all individuals carrying the latter allele from our analyses. The univariate analyses showed that individuals carrying the CCR2-64I allele had a 35% reduced risk of progression to AIDS when compared to wild type homozygous subjects (HR = 0.65, p = 0.015) (Table II). The effect remained significant in the multivariate analysis (HR = 0.7, p = 0.026) (Table II). No significant correlation was detected between the polymorphism CCR5-59029A and the course of infection, even in the absence of the protective alleles CCR5-∆32 and CCR2-64I (Table II). The SDF1-3'A allele also failed to show any association with changes in the HIV disease progression in our casuistic when analysed individually (Table II). We also performed an expectation-maximisation analysis of the polymorphisms clustered at chromosome 3p21 and the results did not differ (Supplementary data). We further analysed the effect of different allele combinations among those studied. Subjects were grouped in the following categories according to the CCR2 and SDF1 genotypes: (i) SDF1-+/+ and CCR2-+/+ (n = 109), (ii) SDF1-+/3'A or SDF1-3'A /3'A and CCR2-+/+ (n = 35), (iii) SDF1-+/+ and CCR2-+/64I or CCR2-64I/64I (n = 72) and (iv) SDF1-+/3'A or SDF1-3'A/3'A and CCR2-+/64I or CCR2-64I/64I (n = 15). Cox regression analyses were conducted and the HR was determined when comparing category 1 (wild-type homozygous) with the remaining three categories. Subjects harbouring SDF1-3'A in the absence of CCR2-64I (category 2) had a 15% reduction in the risk of progression to AIDS (HR = 0.85, CI 0.65-1.1) (Table III), while subjects carrying CCR2-64I but not SDF1-3'A (category 3) had a 26% reduction in the risk for the same outcome (HR = 0.74, CI 0.5-1.11) (Table III). Subjects carrying both alleles had a 53% reduction in the risk of progression to AIDS (HR = 0.47, CI 0.27-0.83, p = 0.03) (Table III). There was no significant correlation between these combined genotypes and decline of CD4+ T-cell counts below 200 cells/µL or time to death (Table III). Subjects harbouring at least one variant allele (CCR5-∆32, CCR2-64I or SDF1-3'A; n = 131) displayed a significant reduction in the risk of progression to AIDS (HR = 0.71, p = 0.03) when compared to those with wild-type genotypes. Survival curves were constructed for both groups (Figure 1) and a significant difference was found (p = 0.01, log-rank test). No difference between groups was found in the risk of the decline of CD4+ T-cell counts to < 200 cells/µL or death. Discussion This study describes the genotype and allele frequencies of the polymorphisms CCR5-∆32, CCR2-64I, CCR5-59029A and SDF1-3'A in an HIV-1-infected population from Southern Brazil. Most importantly, however, this study is the first to be conducted in Brazil that investigates the effects of such polymorphisms on the clinical course of HIV infection. The polymorphisms analysed in this study exhibit variable frequencies in distinct populations (Smith et al. 1997, McDermott et al. 1998, Su et al. 1999, Gonzalez et al. 2001). As mentioned previously, the Brazilian population has a complex genetic background (Callegari-Jacques et al. 2003, Pena 2005). A study analysing different populations in Brazil showed the predominance of three mitochondrial lineages. The relative frequency of the lineages varied considerably among Brazilian regions. The majority of lineages from the North have Amerindian origin (54%), whereas African ancestry is more common in the Northeast (44%) and European is more common in the South (66%). The South-eastern Region presented a balance between the frequencies of the three main lineages (Pena 2005). This genetic diversity plays an important role in the variable frequencies of these polymorphisms found within the country. The CCR5-∆32 allele has been found in Brazilian populations in frequencies between 2.2-6.8% (Munerato et al. 2003, Vargas et al. 2006, Reiche et al. 2008, Rigatto et al. 2008). Here, we found a frequency of 2.4%. Similarly, in another study conducted in Southern Brazil, the CCR5-∆32 frequency was 2.9% among HIV-1 seropositive patients with AIDS, 5.6% among those seropositive and asymptomatic, 5.6% in healthy blood donors and 5.51% in HIV-1-exposed seronegative subjects. CCR5-Δ32 allelic frequencies were not different when the self-reported racial characteristics of the individuals evaluated were considered (Reiche et al. 2008). This allele has not been found among South American native Indians, corroborating the hypothesis of a European origin of this allele and its introduction to the continent through migration (Leboute et al. 1999). It is known that the distribution of the CCR2-64I mutation and its allelic frequency varies based on the population and ethnic groups studied (Martinson et al. 2000). Acosta et al. (2003) investigated the CCR2-64I mutation in three different Brazilian populations and found it at a frequency of 26% in Amerindians in the North, 30% among German descendants in the South and 14% in African descendants in the Northeast. In this study, we found the CCR2-64I allele to have a frequency of 11.3%. Another study in the Southeast found an allele frequency of 8.15%. These data are similar to those found in North America and Western Europe (Munerato et al. 2003). The SDF1-3'A allele has a wide global prevalence. It has been found in high frequencies in Oceania and in lower frequencies in Africa (Su et al. 2000). In Brazil, Grimaldi et al. (2010), studying the same three populations that were analysed by Acosta et al. (2003) for CCR2-64I, found an allelic frequency of 21% in the population of the South, similar to the frequency found in this study (20.1%). In the Northeast, the frequency was found to be 17% and, in the North, frequencies of 24% and 5% were observed in two different Amerindian tribes. Finally, the CCR5-59029A allele has been found in frequencies above 40% in different populations (McDermott et al. 1998, O'Brien et al. 2000). In agreement with these results, the frequency found in this study was 48.7%. Similarly, a study conducted in South-eastern Brazil found a frequency of 44.2% in HIV-infected individuals (Rigatto et al. 2008). In our study, the univariate analysis showed that the presence of CCR5-∆32 conferred significant protection from the decline of CD4+ T-cell counts below 200 cells/µL in accordance to previous study (Eugen-Olsen et al. 1997). The slower progression observed in CCR5-∆32 heterozygous subjects could be attributed to the reduced cell surface expression of CCR5 (Wu et al. 1997), the main co-receptor for R5 strains of HIV-1, which predominate in the early stages of infection (Shankarappa et al. 1999). The influence of the CCR5-Δ32 allele on the risk for CD4+ T-cell count decline lost its significance when the model was adjusted for baseline HIV viral load, CD4+ counts and age. This result suggests that, despite its protective effect, the CCR5-∆32 genotype alone does not provide additional information on prognosis when other laboratorial markers are known, as suggested previously (Taylor et al. 2000). However, Kulkarni et al. (2008) showed that testing for possession of the CCR5-∆32 allele in association with determining the copy number of CCL3L1, a potent CCR5 ligand, is predictive of disease progression independent of laboratory markers. This study illustrates how genomic information might be applied to achieve practical benefits (Kulkarni et al. 2008). In contrast to the protective effect of the CCR5-∆32 allele in the early stages of infection, we found the polymorphism to be correlated with an accelerated risk of death after AIDS diagnosis. This finding is congruent with data by Brumme et al. (2005), showing that individuals heterozygous for CCR5-∆32 present a 2.5X increased risk of harbouring CXCR4-tropic HIV strains compared to homozygotes for the wild-type CCR5 allele. Considering that the emergence of X4 strains is usually associated with decreases in CD4+ T-cell counts and with clinical progression (Connor et al. 1997), this emergence could explain the increased risk of progression to death in the late stages of infection found herein. Several studies have suggested that the allele CCR5-59029A is associated with an accelerated disease progression (McDermott et al. 1998, An et al. 2000, Kaur et al. 2007). In our study, such correlation was not evidenced. Martin et al. (1998) and An et al. (2000) suggested that the effect of this polymorphism is more evident in the first two-four years after seroconversion and subtler or even absent later on the infection. Because our study was conducted using chronically infected subjects, it is possible that this fact contributed to the lack of correlation observed. In our study, individuals harbouring the CCR2-64I polymorphism showed a reduced risk for progression to AIDS. Nevertheless, as also reported by Kostrikis et al. (1998), the presence of this polymorphism did not alter the risk of CD4+ T-cell decline. The mechanism by which CCR2-64I confers protection from disease progression remains unknown. The CCR2 receptor acts as a minor co-receptor for HIV-1 and the polymorphism CCR2-64I did not alter its properties as a chemokine receptor or as an HIV co-receptor (Lee et al. 1998). Some studies have suggested that this allele might diminish the expression of CCR5 and CXCR4 on the cell surface (Kostrikis et al. 1998, Mellado et al. 1999, Nakayama et al. 2004). SDF1-3'A was also correlated with protection to disease progression in our study, but this effect was influenced by the CCR2 genotype. SDF1-3'A-harbouring individuals were only protected in comparison to those carrying the wild-type form of both SDF1 and CCR2 genes. The considerable number of subjects harbouring CCR2-64I combined with the wild-type form of SDF1 (n = 30) likely masked the protective effect of the SDF1-3'A allele. However, subjects carrying both polymorphisms showed a 53% reduction in the risk of AIDS progression. This protection was higher than that observed in subjects carrying only one of those alleles, suggesting a cumulative effect. Our results highlight the need for analysing polymorphisms in genes that share a common molecular pathway in a conjoined manner. The protective effect of the CCR5-∆32, CCR2-64I and SDF1-3'A alleles was confirmed when comparing subjects carrying any of those polymorphisms to subjects who were homozygous for the wild-type form of all three genes. The reduction in risk for disease progression observed in individuals with these mutations provides evidence for the role of such polymorphisms and therefore the genetic background of the population, on HIV disease progression. Some studies have investigated the role of polymorphisms in chemokine and chemokine receptor genes during the use of antiretroviral (ARV) therapy (O'Brien et al. 2000, Brumme et al. 2001, Passam et al. 2005). Our results showed that the use of ARV therapy did not influence the protective effects seen in the overall casuistic (Table II), as similar HR values were observed. It is noteworthy to mention that the present study carries some important limitations. One of them is the reduced number of subjects included in the study, which may have masked some correlations among those analysed. Another limitation refers to the analysis of a chronically infected cohort, which may again mask some of the effects, as discussed above. However, a meta-analysis of the polymorphisms studied here has shown that similar results are obtained when analysing either recently or chronically-infected subjects (Ioannidis et al. 2001), suggesting that our results are relevant, particularly in this poorly studied population. In summary, we demonstrated the protective effect of the CCR5-∆32, CCR2-64I and SDF1-3'A alleles on the risk for AIDS progression in a Brazilian population. An important interaction between the alleles CCR2-64I and SDF1-3'A was observed. A temporal effect relative to the time of seroconversion was shown for the CCR5-∆32 allele. Altogether, our results highlight the importance of studying genetic polymorphisms in different populations and their potential use as prognostic markers for HIV disease progression. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11071t1.jpg] [oc11071f1.jpg] [oc11071t2.jpg] [oc11071s1.jpg] [oc11071t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}