 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 3, 2011, pp. 467-474 ARTICLES Neutralising antibodies for West Nile virus in horses from Brazilian Pantanal Alex Pauvolid-CorrêaI, +; Maria Alejandra MoralesII; Silvana LevisII; Luis Tadeu Moraes FigueiredoIV; Dinair Couto-LimaI; Zilca CamposIII; Marcia Furlan NogueiraIII; Edson Elias da SilvaV; Rita Maria Ribeiro NogueiraI; Hermann Gonçalves SchatzmayrI, * I Laboratório de Flavivírus, Instituto Oswaldo Cruz-Fiocruz, Ministério da Saúde, Avenida Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil * In memoriam Received 16 December
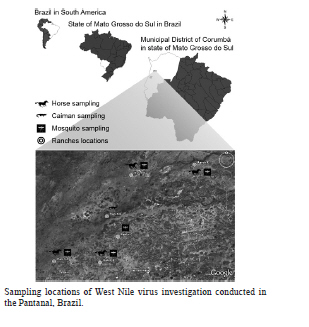
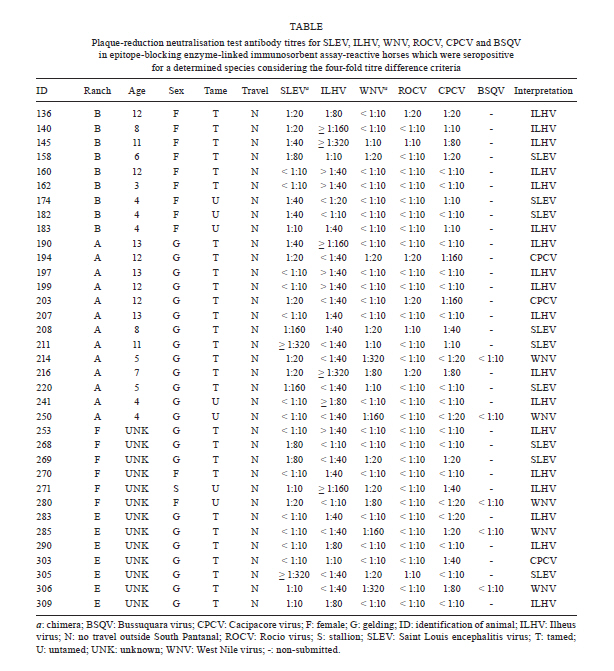
2010 Code Number: oc11080 Abstract Despite evidence of West Nile virus (WNV) activity in Colombia, Venezuela and Argentina, this virus has not been reported in most South American countries. In February 2009, we commenced an investigation for WNV in mosquitoes, horses and caimans from the Pantanal, Central-West Brazil. The sera of 168 horses and 30 caimans were initially tested using a flaviviruses-specific epitope-blocking enzyme-linked immunosorbent assay (blocking ELISA) for the detection of flavivirus-reactive antibodies. The seropositive samples were further tested using a plaque-reduction neutralisation test (PRNT90) for WNV and its most closely-related flaviviruses that circulate in Brazil to confirm the detection of specific virus-neutralising antibodies. Of the 93 (55.4%) blocking ELISA-seropositive horse serum samples, five (3%) were seropositive for WNV, nine (5.4%) were seropositive for St. Louis encephalitis virus, 18 (10.7%) were seropositive for Ilheus virus, three (1.8%) were seropositive for Cacipacore virus and none were seropositive for Rocio virus using PRNT90, with a criteria of > four-fold antibody titre difference. All caimans were negative for flaviviruses-specific antibodies using the blocking ELISA. No virus genome was detected from caiman blood or mosquito samples. The present study is the first report of confirmed serological evidence of WNV activity in Brazil. Key words: West Nile virus - Pantanal - neutralisation - equines - caimans - BrazilFinancial support: CNPq, FAPERJ, FIOCRUZ In 1937, an unknown neurotropic virus called West Nile virus (WNV) was isolated from the blood of a low-grade febrile woman from northeastern Uganda (Smithburn et al. 1940). Afterwards, WNV was detected in Africa, the Middle East, Asia and Europe and was associated mainly with sporadic or size-limited outbreaks of WNV fever, which was usually a self-limited and uncomplicated febrile syndrome (Couissinier-Paris 2006). However, in the mid-1990s, a major WNV fever epidemic in Europe with a high rate of neurological infections occurred in the lower Danube valley and Bucharest areas in Romania (Tsai et al. 1998). Concomitantly, human and animal outbreaks of WNV fever with neurological disorders were reported in the old world, which warned of the change of WNV fever impact on animal and public health (Tber Abdelhaq 1996, Platonov et al. 2001). In 1999, WNV definitively reemerged as a worldwide major public health and veterinarian concern when it was detected in the Western Hemisphere for the first time as the cause of human and equine encephalitis outbreaks concomitant with extensive mortality in birds in the northeastern United States of America (USA) (CDC 1999). The virus rapidly spread through North America and towards Latin America (Komar & Clark 2006). There have been recent reports of serological evidence of WNV activity in Colombia (Mattar et al. 2005), Venezuela (Bosch et al. 2007) and especially Argentina, where anti-WNV antibodies have been detected in birds (Diaz et al. 2008) and the virus was isolated for the first time in South America from the brains of horses that had died of encephalitis (Morales et al. 2006). However, most South American countries, including the largest, Brazil, remain without reports of WNV infection. Alerted by the periodical outbreaks of WNV encephalitis in the USA (Lindsey et al. 2010) and the serological evidence of WNV activity in South America, we performed this study. The Brazilian Pantanal was selected as a suitable place for the study of WNV circulation because of the following factors: (i) natural foci of WNV infections have been mostly described in wetland ecosystems, such deltas and flood plains, as reported in France (Joubert et al. 1970), Venezuela (Bosch et al. 2007), Romania (Tsai et al. 1998) and Italy (Autorino et al. 2002), (ii) the local movement of migratory birds (Nunes & Tomas 2008), (iii) the unsuccessful efforts of the detection of WNV-specific antibodies in the free-ranging birds in Brazilian coastal areas (MS 2003, 2004), (iv) the recent detection of arbovirus vectors blood-feeding on human and equine hosts in the area (Pauvolid-Corrêa et al. 2010a) and, finally, (v) the serological evidence of arbovirus activity in the area (Iversson et al. 1993), which includes St. Louis encephalitis virus (SLEV) (Pauvolid-Corrêa et al. 2010b), a species that is antigenically related to WNV (Smithburn 1942). In view of these data, the hypothesis that the Brazilian Pantanal is one of the most susceptible areas to WNV circulation in Brazil (Pauvolid-Corrêa & Varella 2008) seemed promising. The Pantanal is a tropical, seasonal wetland of approximately 140,000 km2 that is fed by tributaries of the upper Paraguay River in the centre of South America, which covers mainly Brazilian but also Paraguayan and Bolivian territories and is classified as one of the largest freshwater wetland ecosystems in the world (Alho 2005). The Brazilian Pantanal, which represents 85% of the total Pantanal area, is located within the states of Mato Grosso do Sul and Mato Grosso in Central-West Brazil, which are recognised as South and North Pantanal, respectively (Alho et al. 1988). This region is ecologically classified into sub-regions according to vegetation, flooding and physiography (Silva & Abdon 1988). The Nhecolândia sub-region that is situated in South Pantanal is one of the largest, comprising approximately one fifth of the total area, and is characterised by hundreds of shallow lakes that display varying degrees of salinity and coalescence with the system during floods (Adámoli 1982). Recent studies have demonstrated that some crocodilian species may be infected by WNV (Steinman et al. 2003, Jacobson et al. 2005) and that Alligator mississippiensis may also have a viremic load that affords the infection of blood-feeding Culicidae vectors, which suggests that in areas with high population densities of these alligators, juvenile individuals could play an important role in WNV transmission (Klenk et al. 2004). Taking these data into account, the unsuccessful efforts to detect WNV circulation in the avian hosts in Brazil and the widespread, high prevalence of the Crocodilia species, Caiman crocodilus yacare, in the Brazilian Pantanal (Campos et al. 2005), our strategy was to investigate the WNV circulation in the potential vectors, dead-end hosts and natural secondary amplifying hosts through the collection and testing of mosquitoes and serum samples of horses and wild caimans from the Nhecolândia sub-region. No morbidity was observed during the sampling period and only apparently healthy horses and caimans were sampled in the present study. Materials and Methods Study area - In February 2009, mosquitoes and blood samples from horses and wild caimans were collected in six different cattle ranches of the Nhecolândia sub-region (18º20' 19º40'S and 57º14' 55º00'W) during the rainy season. Considering the free circulation of mosquitoes, horses and wild caimans in large ranches, sample collections for the study were undertaken in a 700-square-kilometre (70,000 ha) area (Figure). The collections for this study were authorised by the Brazilian Institute of Environment and Natural Resources (license IBAMA 18363-1/2009). Samples collections - Adult mosquitoes were captured at sites that were randomly selected using CDC automatic light traps and manual aspirators while landing to blood-feed on horses and research team members as routinely reported. Living mosquito specimens were transported to a field laboratory where they were immobilised by chilling and the species were identified through direct observation of the morphological characteristics according to the taxonomic keys (Lane 1953, Consoli & Lourenço-de-Oliveira 1994). The mosquitoes were grouped into pools of up to 50 individuals that were sorted with respect to trap, site of capture and species and they were then stored in liquid nitrogen in the field. Caimans were captured from sites where a high concentration of these animals was observed. The animals were seized by boat in lentic systems that were formed by intermittent rivers and taken to the banks. All animals were weighed, measured, gendered, numbered and bled before release. The whole procedure was performed only with physical restraint and each animal was worked and released within 15 min. Caiman blood samples were obtained by puncture of the internal jugular vein between the 1st and 2nd cervical vertebrae as described previously (Olson et al. 1975). Blood samples were taken from horses of different ages from different ranches. Concomitantly, an interview was conducted with the rural workers of the ranches that were visited who were asked if they had seen horses with any abnormality, such as clinical signs that involved the central or peripheral nervous system. We also collected information about the sex, age and breed of the horses that were present. Additional information included whether the horse had recently received vaccine or medical treatment and whether any horse had been moved into or out of South Pantanal. Epitope-blocking enzyme-linked immunosorbent assay (blocking ELISA) - Caiman and horse sera were first screened for flaviviruses-reactive antibodies using a blocking ELISA as described previously (Blitvich et al. 2003a, c). Briefly, the ability of the sera to block the binding of the monoclonal antibody 6B6C-1 to the WNV antigen was compared to the blocking ability of negative control horse serum. The monoclonal antibody 6B6C-1 is appropriate to screen for any flavivirus-reactive antibody. Data are expressed as relative percentages. Samples were considered seropositive when the inhibition values produced were > 35%. Plaque-reduction neutralisation test (PRNT) - The seropositive sera of the blocking ELISA was sequentially heat-inactivated and submitted to a PRNT90 as described previously (Russell et al. 1967). Briefly, two-fold dilutions that ranged from 1:10-1:320 of each blocking ELISA seropositive sample were tested for their ability to neutralise plaque formation by the chimeras SLEV and WNV that were derived from highly attenuated Yellow fever virus that contain the premembrane-envelope (prM-E) and prM-E and envelope protein genes, respectively (Arroyo et al. 2001, Pugachev et al. 2004, Komar et al. 2009), and Cacipacore virus (CPCV), Ilheus virus (ILHV) and Rocio virus (ROCV), which are the Brazilian flaviviruses that are phylogenetically related to WNV (Kuno et al. 1998). Serum was considered seropositive to a virus species when it reduced at least 90% of the formation of plaques of this virus at > 1:40 dilutions and its neutralising antibody titre was > four-fold greater than what was observed for the other tested flaviviruses. The WNV seropositive samples were also submitted to PRNT90 for Bussuquara virus (BSQV). Virus isolation - The caiman sera and mosquito samples were assayed for virus isolation. The caiman serum samples at 1:5 dilutions were inoculated into VERO cell culture flasks (Stim 1969). The macerated mosquito pool suspensions were inoculated into C6/36 cell culture flasks (White 1987). The incubated cell flasks were observed during 14 days to look for any evidence of viral cytopathic effect (CPE). Reverse-transcriptase polymerase chain reaction (RT-PCR) - The molecular approach was performed in all caiman serum samples and the mosquito samples in which the inoculated flasks showed CPE in virus isolation. These samples were submitted to RNA extraction using a commercial kit (QIAamp© Viral RNA Mini Kit QIAGEN, USA) in accordance with the manufacturer's instructions and was followed by a generic nested RT-PCR, which sequenced for the detection and identification of arbovirus members of the Flavivirus and Alphavirus genus, as described previously (Sánchez-Seco et al. 2001, 2005). Results Serum samples of 168 horses from four of the six studied cattle ranches were sampled. Most of these equines were adult, tame, healthy thoroughbreds of the Cavalo Pantaneiro breed. All 168 horse sera were first tested using the blocking ELISA and 93 (55.4%) were seropositive. When this subset of 93 seropositive samples was tested using PRNT90, five (3%) were seropositive for WNV, nine (5.4%) were seropositive for SLEV, 18 (10.7%) were seropositive for ILHV, three (1.8%) were seropositive for CPCV, but none were seropositive for ROCV, using the criteria of a > four-fold antibody titre difference among the five flaviviruses that were assayed. All of the WNV-seropositive samples were < 1:10 for BSQV (Table). From the remaining 58 (34.5%) blocking ELISA seropositive samples, 56 (33.3%) samples were considered seropositive for an undetermined flavivirus using the < four-fold antibody titre difference and two (1.2%) were considered seronegative with titres < 1:10 for all of the flaviviruses that were assayed by PRNT90. From the five horses that were seropositive for WNV, two horses had dates of birth available and were four and five years old at the time of blood collection. All five seropositive animals had no history of journey outside South Pantanal or WNV vaccination. All WNV-seropositive samples were retested for confirmation and the antibody serum titres were 1:80 (n = 1), 1:160 (n = 2) and > 1:320 (n = 2). The sera of these two horses with antibody serum titres > 1:320 were retested with higher dilutions and an antibody serum titre of 1:320 was confirmed. These five WNV-seropositive horses came from three out of the four ranches where horses were sampled. Concerning SLEV of the nine seropositive horses from the four ranches sampled, six had dates of birth available with ages that ranged from four-11 years old. All of the SLEV-seropositive horses had never left South Pantanal and their antibody serum titres ranged from 1:40-> 1:320. Of 18 ILHV-seropositive horses, 12 had available dates of birth with ages that ranged from three-13 years old in animals from all of the ranches where horses were sampled. None of these animals had a history of travel and their antibody serum titres ranged from 1:40-> 1:320. From the three horses that were seropositive for CPCV, two had available dates of birth and were 12 years old at the time of blood collection. These two animals were from the same ranch, had never left the region and had antibody serum titres for CPCV of 1:160. Most of the blocking ELISA-seropositive samples had antibody titers < 1:10 for ROCV. Of the samples with antibody titres > 1:10 for ROCV, only one had a titre > 1:40. Because the antibody titre difference for the other flaviviruses assayed was less than four-fold, it was considered seropositive for an undetermined flavivirus. Of the 93 blocking ELISA-seropositive horses, 56 (60.2%) with titre differences less than four-fold were considered seropositive for an undetermined flavivirus. The two blocking ELISA-seropositive samples that had serum titres < 1:10 for all five flaviviruses assayed were considered as seronegative in the present study. Thirty caiman specimens up to 1.5 m in length were captured from two distinct sites of the same ranch. The sera from the 30 caimans displayed negative results to virus isolation, RT-PCR and blocking ELISA and were subsequently not submitted to PRNT90 methods. A total of 1,204 mosquitoes from 10 species were captured at 19 sites that were randomly selected on five ranches. Specimens of the following species were captured: Ochlerotatus scapularis, Psorophora albigenu, Sabethes albiprivus, Culex declarator, Culex quinquefasciatus, Mansonia titillans, Anopheles albitarsis s.l., Psorophora ferox, Anopheles triannulatus s.l. and Psorophora ciliata. The mosquitoes were grouped into 65 pools of up to 50 individuals that were sorted with respect to trap, site of capture and species. Of a total of the 65 mosquito pools that were assayed, one (1.5%) pool with 22 specimens of O. scapularis exhibited CPE four days post-infection (dpi). The same CPE was observed on subsequent passages by inoculation into new C6/36 cell culture flasks four dpi. Nevertheless, despite the CPE, the pool and the supernatant from the sequential passages was negative to flaviviruses and alphaviruses as measured by RT-PCR. Discussion Flaviviruses serological test result interpretation is sometimes complex and requires careful evaluation (Komar 2001), especially where data from areas of intense flaviviruses co-circulation is expected, such as for South American countries. Considering the complex cross-reactivity of flaviviruses, several studies have adopted ELISA as a screening method followed by more virus-specific serological tests, such as PRNT methods (Blitvich et al. 2003b, Morales-Betoulle et al. 2006, Diaz et al. 2008). In the present study, the use of blocking ELISA as a screening method followed by PRNT90 for the most related sylvatic Brazilian flaviviruses allowed for the detection of specific neutralising antibodies to WNV in 3% of the horses tested from the Pantanal region for the first time in Brazil. Based on the results that were obtained with the blocking ELISA, 93 horse sera presented antibodies to flaviviruses. When the same sera were tested using PRNT90 for SLEV, WNV, ILHV, ROCV and CPCV, except for two samples, none had a serum titre < 1:10 for all of the flaviviruses that were tested, which disclosed the high specificity of the blocking ELISA method. According to our data, in conjunction with the fact that blocking ELISA does not require the use of multiple capture antibodies, which designated this test as useful for virtually any vertebrate species, the present study corroborated the application of blocking ELISA as an efficient screening method for flaviviruses serosurveys. We have searched for the circulation of WNV in a country with an intense circulation of other flaviviruses. Therefore, to avoid misinterpretation based on serum cross-reactions it was necessary to employ a maximum specificity PRNT and only consider seropositive samples that promote a > 90% reduction in the number of WNV plaques as has been described previously in South and Central American serosurveys (Quirin et al. 2004, Mattar et al. 2005). Besides SLEV, ROCV, ILHV and CPCV, other Brazilian sylvatic flaviviruses that are related to WNV include the BSQV and Iguape virus (IGUV) (Kuno et al. 1998). Despite the geographic dispersal of BSQV, which is mainly located in the Caribbean and Central America (Gubler et al. 2007), the WNV seropositive samples were also submitted to PRNT90 for BSQV to minimise potential high-titered cross reaction antibodies. However, considering that IGUV was only isolated from sentinel mice in the Southeastern Brazil (Coimbra et al. 1993) and that serosurveys suggest infrequent occurrence of overt infection, in the present study IGUV was not included in the PRNT methods. To minimise the cross-reaction results in this serosurvey to WNV, we considered the seropositivity for WNV and for each of the other flaviviruses assayed, only sera with a 1:40 monotypic reaction or with an antibody titre higher than 1:40 and a four-fold higher to a species than its other related flaviviruses that were assayed. However, considering these criteria, 56 (60.2%) of the 93 blocking ELISA-seropositive samples were seropositive for an undetermined flavivirus species. The high prevalence of these inconclusive results may be attributed to the cross-reacting antibodies that are usually observed among species of a Japanese encephalitis virus group (JEVG) (Tarr & Hammon 1974) and to the original antigenic sin phenomenon, where some individuals that are sequentially infected by a heterologous flavivirus species can boost antibody levels against the original virus, which produces heterologous cross-reacting antibodies (Inouye et al. 1984). Considering the similar maintenance cycles of the flaviviruses that were assayed together with the hypothesis of its co-circulation in the South Pantanal, the original antigenic sin phenomenon may have had an effect on the difference of antibody titres among the flaviviruses that were assayed and some sequential infections by a heterologous flavivirus species may not have been detected. Of the 56 horses that were seropositive for an undetermined flavivirus species, 13 (23.2%) had antibody titres > 1:40 for WNV, nine (69.2%) of which had antibody titres > 1:80. Therefore, it is possible that the other 13 (7.7%) of the 168 horses that were tested had contact with WNV. It is noteworthy that neutralising antibody titres are frequently more than two-fold lower in PRNT with chimeras WNV and SLEV than with wild virus (Pugachev et al. 2004, Komar et al. 2009). Therefore, it is also quite possible that some low titre WNV and SLEV-seropositive samples were not detected. The detection of specific neutralising antibodies to WNV in 3% of horses tested in the Pantanal region from Brazil was reported in this study. This prevalence is similar to the values previously reported for horses from other countries of Central and South America, such as Venezuela, where a prevalence of 4.3% was described (Bosch et al. 2007); Colombia, with a prevalence of 9% (Mattar et al. 2005), and El Salvador, with the highest seropositivity to WNV of 13.7% (Cruz et al. 2005). Despite the adoption of the same criteria for the detection of WNV-seropositive animals, it is important to note the difference of the flaviviruses that were used in the PRNT methods that were performed in different areas. The CPCV, which is a Brazilian virus of JEVG, is as phylogenetically related to WNV as SLEV and is even more closely related to WNV than ILHV, BSQV or ROCV, which belong to other flavivirus serocomplexes (Kuno et al. 1998). Although CPCV is a non-pathogenic arbovirus to date, we detected many samples with neutralising antibodies for both CPCV and WNV, which suggest that in future serosurveys for WNV and other sylvatic flaviviruses to be conducted in Brazil, the investigation for CPCV should be included as a differential diagnosis. The highest seropositivity for ILHV was expected because the circulation of ILHV in South Pantanal horses was reported in the 1990s (Iversson et al. 1993). Although ROCV, which is an indigenous Brazilian flavivirus species, was the causative agent of an extensive encephalitis epidemic in Southeastern Brazil in the 1970s (Tiriba et al. 1976), the detection of neutralizing antibodies for ROCV nowadays is unusual. Our findings corroborate this data because except for one sample, all blocking ELISA-seropositive horses had PRNT antibody titres < 1:40 for ROCV. One should be aware that, although phylogenetically related to ILHV (Medeiros et al. 2007) that presented the highest prevalence in our study, none of the horses tested were seropositive for ROCV, which demonstrates the high-quality specificity of the PRNT assay. Based on our serological data, we reported indirect evidence that argues for the silent activity of WNV in the Brazilian Pantanal. Considering this hypothesis and taking into account the age available for the seropositive animals of four and five years old, we can infer that at least during the period from 2005-2009, WNV circulated in the area. The detection of SLEV-seropositive four-year-old horses suggested that some were infected by SLEV between 2005-2009, which corroborates the recent report of the detection of neutralising antibodies to SLEV in horses sampled in 2007 in the same area of the present study (Pauvolid-Corrêa et al. 2010b). For CPCV, the two seropositive horses with available dates of birth were 12 years old; therefore, the period of probable circulation was difficult to determine. The circulation of ILHV in South Pantanal was described in the 1990s (Iversson et al. 1993) and this putative circulation has remained. However, according to the age of our 18 ILHV-seropositive animals, we can only deduce that between 2006-2009, ILHV circulated in the area. Although a better estimation of recent circulation of these flaviviruses for future surveillance should be based on IgM assays, our data suggested the viral activity after 2005 of at least three flaviviruses, including WNV, in the Brazilian Pantanal. We encourage further more encompassing surveys for WNV in the South Pantanal and other bordering areas that especially focus on virus isolation to definitively confirm the circulation of WNV in the region, and consequently, identify vectors and vertebrate hosts that are involved in the virus local maintenance cycle and transmission. Despite of the serological evidence of horse infections by WNV in the Pantanal region presented here, morbidity was not observed, which suggests that WNV could be circulating through silent enzootic cycles in an area with pristine conditions. In fact, WNV has not been related to horse disease in most of the South American countries with the exception of Argentina (Morales et al. 2006). However, WNV is an important cause of encephalitis in horses in the USA and should be considered in the future in the differential diagnosis of neurological infections in horses from South Pantanal. Despite of the small number of caimans that were sampled, the negative results obtained using virus isolation, RT-PCR and blocking ELISA suggested that the participation of caimans in flaviviruses cycles in South Pantanal is improbable. Nevertheless, the high prevalence of this crocodilian species in the Pantanal and the high concentration that was observed in some remaining collections of water during the dry season associated to the present data of serological evidence of the activity of members of JEVG in the area justify the need of further arbovirus investigations in local caimans. All mosquito species identified in the present study were previously reported in the Nhecolândia sub-region (Pauvolid-Corrêa et al. 2010a). Moreover, the non-detection of Flavivirus and Alphavirus RNA in the O. scapularis pool that caused CPE in C6/36 cells implied a virus identity that was not related to WNV, SLEV or alphaviruses, which would have been indicated by the RT-PCR. Further investigations of the other arbovirus groups are currently in progress. Finally, we have reinforced that the Brazilian Pantanal is an interesting scenario where sylvatic flaviviruses cycles are probably maintained in silent progression in a relatively balanced ecosystem. The recently increased environmental degradation of this sensitive, complex biome could lead to changes in the intricate ecological relationships that disturb natural enzootic arbovirus cycles, which potentially favours local arbovirus outbreaks. Acknowledgements To the cordial people of the Pantanal, who have wisely managed to keep the Pantanal preserved through the centuries, especially the rural workers of the ranches sampled including Marcelo Santos, Francisco Rondon, Luis Fernando Santiago, Gilberto Silva, Marcelo Dias, José Mendes, Marcos Moraes, Luiz Graça, Luiz da Silva, Benis da Silva and Sebastião Gonzaga, to Henrique de Jesus, Augusto Silva, Cremilson Gonçalves, Murilo Maciel and Oziel da Silva, of Embrapa Pantanal to Victoria Luppo, Pablo Baroni, Lucas Bogdanic, Stella Fuster and Silvina Coenaga, of Instituto Nacional de Enfermidades Virales Humanas, to Vanessa Muller, of Faculdade de Medicina de Ribeirão Preto, and to Heitor Herrera, Fernando Tavares, Flávia dos Santos, Ana Bispo and Elba Lemos, of Fiocruz, for their contributions. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11080t1.jpg] [oc11080f1.jpg] |
| |||||||||

{kind=link}
{kind=link}