 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 3, 2011, pp. 507-509 SHORT COMMUNICATIONS Miltefosine induces programmed cell death in Leishmania amazonensis promastigotes Fernanda de Aquino MarinhoI; Keyla Cristiny da Silva GonçalvesI; Selma Soares de OliveiraI; Ana-Carolina de Siqueira Couto de OliveiraII; Maria BellioII; Claudia Masini d'Avila-LevyIII; André Luis Souza dos SantosI; Marta Helena BranquinhaI, + IDepartamento
de Microbiologia Geral, Instituto de Microbiologia Prof. Paulo de Góes,
Centro de Ciências da Saúde, Universidade Federal do Rio de Janeiro,
Av. Carlos Chagas Filho 373, 21941-902 Rio de Janeiro, RJ, Brasil Received 29 October
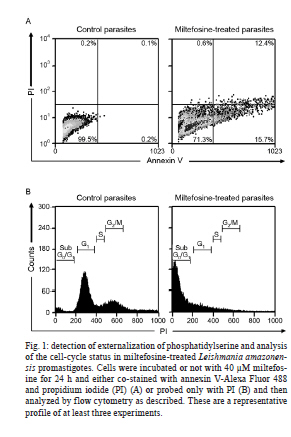
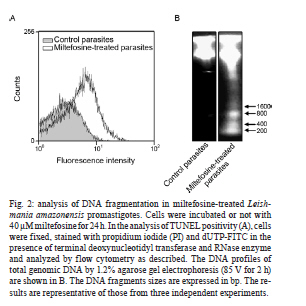
2010 Abstract In the current study, we evaluated the mechanism of action of miltefosine, which is the first effective and safe oral treatment for visceral leishmaniasis, in Leishmania amazonensis promastigotes. Miltefosine induced a process of programmed cell death, which was determined by the externalization of phosphatidylserine, the incorporation of propidium iodide, cell-cycle arrest at the sub-G0/G1 phase and DNA fragmentation into oligonucleosome-sized fragments. Despite the intrinsic variation that is detected in Leishmania spp, our results indicate that miltefosine causes apoptosis-like death in L. amazonensis promastigote cells using a similar process that is observed in Leishmania donovani. Key words: leishmaniasis - chemotherapy - apoptosis Leishmaniasis is a protozoan disease that affects approximately 12 million people worldwide, particularly in tropical and sub-tropical regions. Over 15 species of Leishmania are known to cause the two main clinical manifestations of leishmaniasis, which are cutaneous leishmaniasis (CL) and visceral leishmaniasis (VL). In the New World, CL has increased in the last 20 years in almost all Brazilian states and includes cases that are caused by infection with Leishmania amazonensis. Although parenteral administration of pentavalent antimony organic compounds remains the first choice therapy for all leishmaniasis syndromes, antimony-resistance, low patient compliance and the high frequency of side effects are still relevant problems that are associated with this treatment. In addition, variations in efficacy as a result of intrinsic variations in drug sensitivity limit the arsenal of anti-leishmanial drugs, as each Leishmania sp. has specific biochemical and molecular characteristics (Croft et al. 2006). Miltefosine or hexadecylphosphocholine, which is an alkylphosphocholine ether-lipid analogue that was originally developed for the treatment of cutaneous metastasis from mammary carcinomas, has been shown to be the first effective and safe oral treatment for VL in India (Sundar et al. 2002). Moreover, miltefosine has been successfully used to treat patients with antimony-resistant VL, VL in children and CL in South America (Croft & Engel 2006). However, limited reports are available on the efficacy of miltefosine against VL and CL in other endemic areas or against other Leishmania spp and the leishmanicidal mechanisms of miltefosine. Previous reports showed that there is a significant difference between the IC50 values after 72 h of miltefosine treatment of promastigotes from the species Leishmania donovani, Leishmania major, Leishmania tropica, Leishmania aethiopica, Leishmania mexicana, Leishmania panamensis, Leishmania braziliensis, Leishmania amazonensis and Leishmania infantum. The IC50 values ranged from 0.12-1.32 µM in L. donovani to 4.8-13.1 µM in L. major (Escobar et al. 2002, Santa-Rita et al. 2004, Cabrera-Serra et al. 2007). In addition, the induction of apoptosis following miltefosine treatment has already been reported in L. donovani (Paris et al. 2004, Verma et al. 2007). Our study aimed to evaluate its mode of action against L. amazonensis to delineate the implications of miltefosine treatment in this parasite. In the current study, we showed that the addition of miltefosine to L. amazonensis promastigotes (MHOM/BR/75 Josefa strain) induced a type of death that shared many features with metazoan apoptosis. We employed the same approach used by Paris et al. (2004), in which L. donovani cells were treated with at least 40 µM miltefosine for 24 h to display apoptosis-like changes. The exponential-phase promastigotes (108 cells/mL) were resuspended in Schneider's insect medium that was supplemented with 10% foetal bovine serum, incubated or not with miltefosine (Aeterna Zentaris, Canada) at 40 µM for 24 h and then washed twice with phosphate buffered saline (PBS) (pH 7.2). To demonstrate that the mechanism of cell death that was triggered by miltefosine is through apoptosis and not necrosis, miltefosine-treated promastigotes were double-stained with Alexa Fluor-conjugated annexin V and propidium iodide (PI). The annexin V-binding assay was performed according to the manufacturers' instructions (Invitrogen, USA) and incubated for 20 min in the dark at 20-25ºC. The intensity of Alexa Fluor-conjugated annexin V labelling was recorded using a FACSCalibur flow cytometer and analysed with CellQuest software. The percentage of positive cells was assessed for each histogram. As shown in Fig. 1A, a significant percentage (15.7%) of miltefosine-treated cells stained positive for annexin V and negative for PI compared to the percentage of untreated cells (0.2%). The percentage of cells that were annexin V and PI-positive was 12.4% and the percentage of cells that were only PI-positive was 0.6%. These data suggested that miltefosine induced cell death by apoptosis, which was shown by PI incorporation and annexin V binding. Flow cytometry analysis after PI labelling was performed to verify the number of cells in the sub-G0/G1 phase. Miltefosine-treated promastigotes were fixed in chilled methanol and incubated overnight at -20ºC. After two washes in PBS, promastigotes were resuspended in 0.5 mL of PI (10 µg/mL in PBS) containing RNase A (248 U/mL) and the mixture was incubated for 20 min in the dark at room temperature. The fluorescence intensity of PI was analysed with a FACSCalibur flow cytometer and CellQuest software. The treatment of cells with 40 µM miltefosine for 24 h induced cell-cycle arrest at the sub-G0/G1 phase (Fig. 1B). The proportion of cells was highest in this phase, which suggested that miltefosine induced DNA degradation in promastigotes. To strengthen the hypothesis that miltefosine induced the apoptotic process in L. amazonensis, DNA fragmentation in L. amazonensis (108 cells) was analysed using the terminal deoxynucleotidyl transferase-mediated dUTP Nick End Labelling (TUNEL) assay with the APO-BrdUTM TUNEL Assay Kit according to the manufacturer's instructions (Invitrogen, USA). The qualitative analysis of DNA fragmentation was performed by electrophoresis with a 1.2% agarose gel (85 V for 2 h) using total genomic DNA that was extracted from untreated and miltefosine-treated promastigotes (Rotureau et al. 2005). The treatment of promastigotes with 40 µM miltefosine induced an increase in nuclear DNA fragmentation, which was shown by dUTP-FITC incorporation (Fig. 2A). The results suggest that the antileishmanial effect is mediated by apoptosis. In addition, DNA analysis by agarose gel electrophoresis revealed DNA fragmentation into oligonucleosome-sized fragments (in multiples of 200 bp) in promastigotes that were treated with 40 µM miltefosine for 24 h (Fig. 2B). However, untreated cells did not show any DNA fragmentation (Fig. 2B). The observation of cells that were PI-positive and annexin V-positive (Fig. 1A) and displayed a certain degree of smearing in the DNA (Fig. 2B) after miltefosine treatment indicated the possibility of necrosis-like death in later stages. This result is not surprising because cells at a very advanced stage of apoptosis resemble necrotic cells, which causes difficulties in the ability to discriminate between late apoptotic and necrotic cells (Verma et al. 2007). Mechanisms of programmed cell death in Leishmania spp are activated in response to various chemotherapeutic stimuli such as sodium stibogluconate (Pentostam), amphotericin B and meglumine antimonate (Glucantime), as well as nutrient deprivation (Lee et al. 2002). Apoptosis greatly affects the host-parasite relationship, because the survival of the parasite in the vector and in the host macrophage requires strict control of the parasite population. Apoptosis may be a useful mechanism to avoid the loss of an entire parasite population and subsequently influences the chemotherapeutic strategies to limit the parasite population (Debrabant et al. 2003, Wanderley & Barcinski 2010). Other alkylphospholipids such as edelfosine, ilmofosine and perifosine have been shown to display antiparasitic activity against Leishmania spp in vitro (Escobar et al. 2002, Azzouz et al. 2005). Perifosine and miltefosine exhibited a similar activity against L. amazonensis in vitro, whereas edelfosine showed the lowest activity. However, the opposite profile was detected for other Leishmania spp, such as L. major (Cabrera-Serra et al. 2007). Edelfosine, miltefosine and ilmofosine inhibited the proliferation of intracellular amastigotes of L. amazonensis. The ultrastructural analysis of edelfosine-treated promastigotes and amastigotes indicated the mitochondrion as the main target. However, plasma membrane alterations and autophagic structures were detected in promastigotes (Santa-Rita et al. 2004). The activity of these compounds was evaluated against L. amazonensis using an in vivo mouse model, which showed that perifosine was the most effective compound. Therefore, perifosine is now considered a novel potential treatment against L. amazonensis--mediated CL (Cabrera-Serra et al. 2008). The combined use of several techniques in the current study conclusively showed that L. amazonensis displayed apoptosis-like cell death after miltefosine treatment. Further studies may shed light on possible targets of miltefosine action. Because miltefosine has been in clinical use in recent years as an anti-leishmanial therapy, a better understanding of the mechanisms that regulate cell death may help us to design novel therapeutic strategies against Leishmania parasites. Further studies are required to evaluate the effects of miltefosine on strains that have become resistant to current treatments. Acknowledgements To Norton Heise (Instituto de Biofísica Carlos Chagas Filho, UFRJ), for donating miltefosine. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11087f2.jpg] [oc11087f1.jpg] |
| |||||||||

{kind=link}
{kind=link}