 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
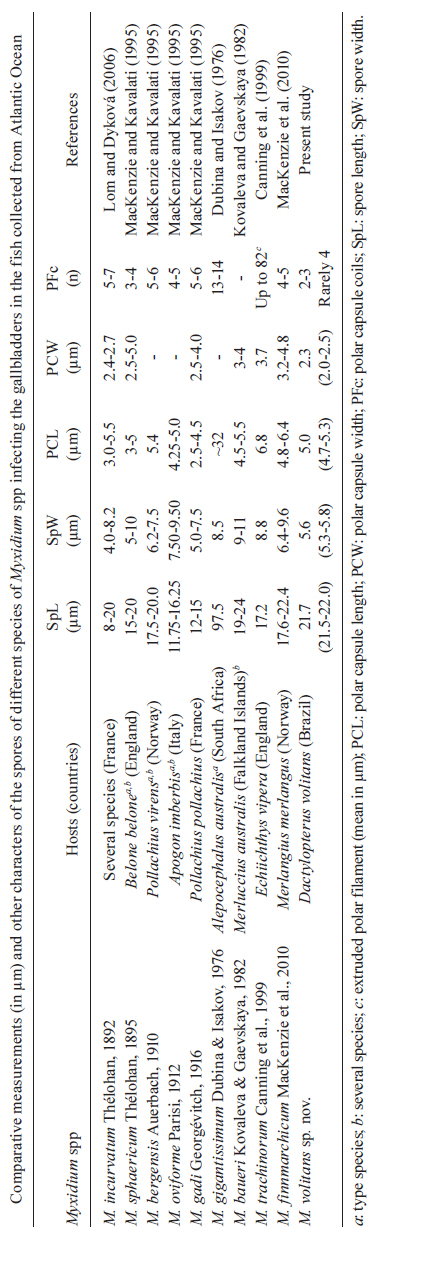
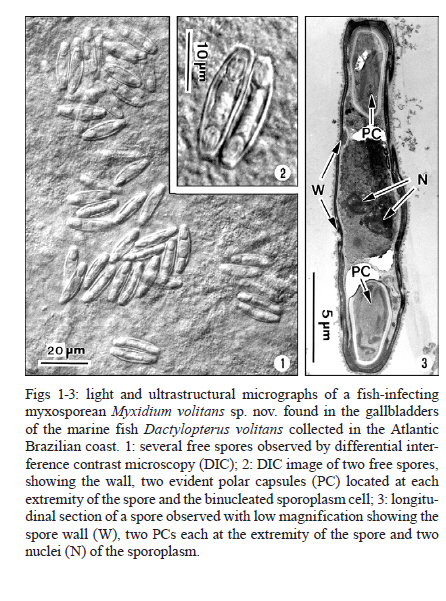
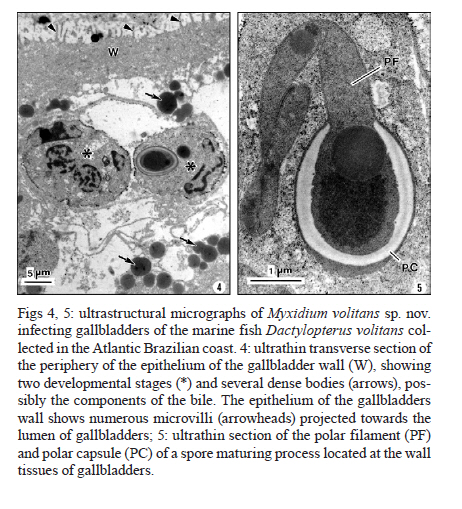
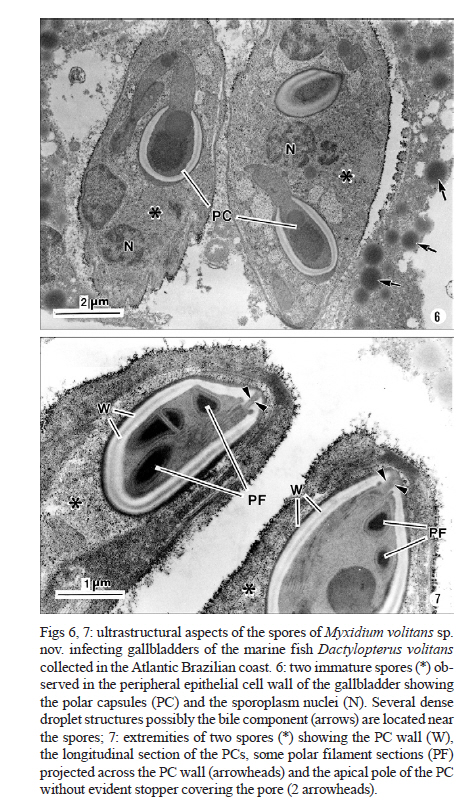
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 4, August, 2011, pp. 557-561 ARTICLES Myxidium volitans sp. nov., a parasite of the gallbladder of the fish, Dactylopterus volitans (Teleostei: Triglidae) from the Brazilian Atlantic coast - Morphology and pathology Carlos AzevedoI, II, +; Graça CasalI, III; Sérgio Carmona São ClementeIV; Leila Maria Silva LopesIV; Patrícia MatosV; Abdel Azeem Abdel-BakiII; Elsa OliveiraI; Edilson MatosVI IDepartamento
de Biologia Celular, Instituto de Ciências Biomédicas e Laboratório
de Patologia, Centro Interdisciplinar de Investigação Marinha
e Ambiental, Universidade do Porto, Largo Prof. Abel Salazar 2, P-4099-003 Porto,
Portugal Financial support: CNPq, CAPES Received 29 December
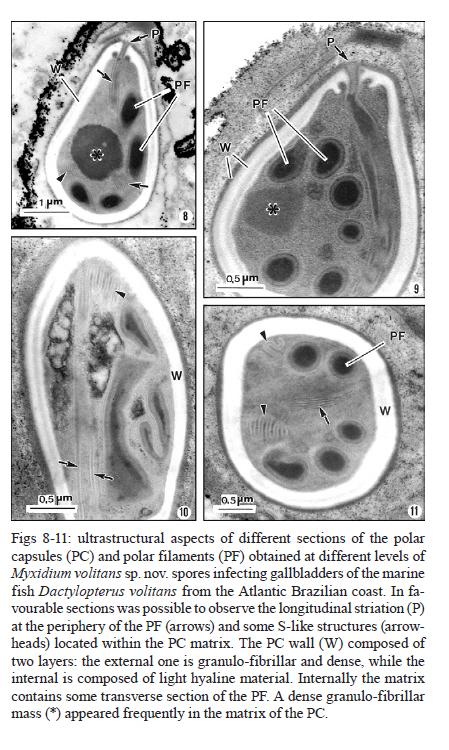
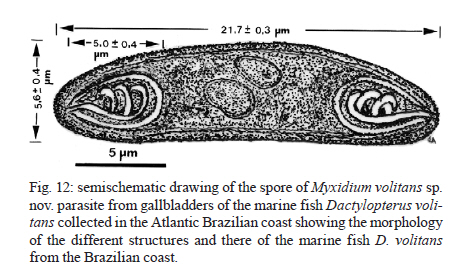
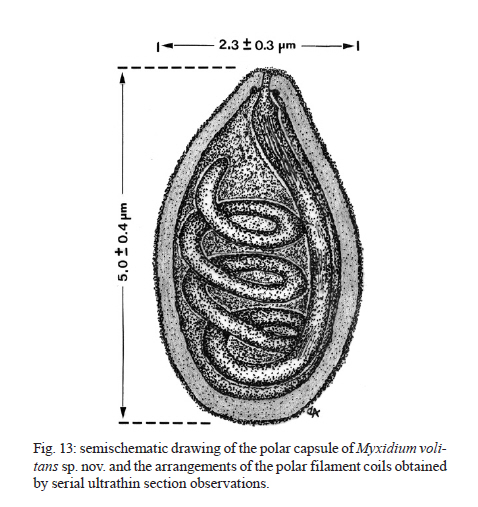
2010 Code Number: oc11095 ABSTRACT Myxidium volitans sp. nov. (Myxozoa: Myxidiidae) parasitizing the hypertrophied green-brownish gallbladder of the teleost Dactylopterus volitans, collected in the Atlantic coast near Niterói, Brazil was described based on ultrastructural studies. The spores were fusiform, sometimes slightly crescent-shaped on average 21.7 ± 0.3 µm (mean ± standard deviation) (n = 50) long and 5.6 ± 0.4 µm (n = 30) wide. The spore wall was thin and smooth, comprising two equally-sized valves joined by a hardly visible sutural ridge. Spores containing two pyriform polar capsules (PC) (5.0 ± 0.4 × 2.3 ± 0.3 µm) (n = 30) are situated in each extremity of the spore. The PC wall was composed of hyaline layer (0.20-0.29 µm thick) and by a thin external granular layer. Each PC contains a polar filament (PF) with irregular arrangements that was projected from its apical region to the bases of PC and coiled laterally from bases to the tip of PC. Some regular striations and S-like structures in the periphery of the PFs with four-five irregular sections were observed. Based on the spore morphology, ultrastructural differences and the specificity of the host we describe this parasite as a new myxosporidian, named M. volitans sp. nov. Key words: Myxozoa - Myxidium volitans sp. nov. - gallbladder - spore - ultrastructure Beside the considerable information available on myxozoan species collected from different geographical areas (Lom & Dyková 2006), little is known about those from South America and in particularly about those from the Brazilian Atlantic coasts, where a diverse assemblage of several hundred species of fish live. Within the myxozoans the genus Myxidium is widely distributed infecting several marine and freshwater fishes, some amphibians and reptiles (Lom & Dyková 2006). Some Myxidium spp infecting the fish galbladders in the Atlantic ichthyofauna from different regions have been recorded (Lom & Dyková 1992, 2006, MacKenzie & Kalavati 1995, MacKenzie et al. 2005): Myxidium incurvatum, collected from several species of the French coasts (Lom & Dyková 1992), Myxidium sphaericum in Belone belone and other hosts, Myxidium bergensis in Pollachius virens and other hosts, Myxidium oviforme in Pagon imberbis from Mediterranean Sea and in other fish species in the Atlantic Ocean, Myxidium gadi in Pollachius pollachius from French region (Mackenzie & Kalavati 1995), Myxidium gigantissimum in Alepocephalus australis off South African coast (Dubina & Isakov 1976), Myxidium baueri in Patagonotothen sima from Falkland Islands (Kovaleva & Gaevskaya 1982, Kalavati et al. 1996), Myxidium trachinarum in Echiichthys vipera from England (Canning et al. 1999) and Myxidium finnmarchicum in Merlangius merlangus from Norway (MacKenzie et al. 2010) (Table). A Brazilian myxosporidian check-list were published (Gioia & Cordeiro 1996) in which the great majority of the results were based on light microscopy, however, this record does not report any Myxidium sp. from the Brazilian coast. Until now, Dactylopterus volitans was not reported as a host of Myxidium sp. This paper describes, based on light and ultrastructural data, a new Myxidium species parasite of a marine fish collected in the Brazilian Atlantic coast. MATERIALS AND METHODS Small fragments of parasitized gallbladder walls and free spores were removed from the bile of the economically important marine fish flying gurnard, D. volitans (Linnaeus, 1758) (Teleostei: Dactylopteridae) (Brazilian common name coió). Sixty four fishes (39 males and 25 females) with an average total length of ~25 cm (range 21-29 cm) (n = 64), collected in the Atlantic coast (22º58'S 43º00'W), near the city of Niterói, state of Rio de Janeiro (RJ), Brazil, were examined and the spores were measured using Nomarski differential interference contrast microscopy equipped with a micrometric ocular and image software. Measurements were based on fresh spores and data are presented as the mean ± standard deviation (range). For transmission electron microscopy (TEM), the free spores and small fragments of parasitized gallbladders were fixed in 5% glutaraldehyde in 0.2 M sodium cacodylate buffer (pH 7.4) for 24 h at 4ºC, washed overnight in the same buffer at 4ºC and post-fixed in 2% osmium tetroxide buffered with the same buffer for 4 h at the same temperature. After dehydration in an ascending graded ethanol series and propylene oxide, the fragments of gallbladders and free spores were embedded in Epon. Semithin sections were stained with toluidine blue. Ultrathin sections, cut with a diamond knife, were double-stained with aqueous uranyl acetate and lead citrate and observed in a JEOL l00CXII TEM operated at 60 Kv. RESULTS During a parasitological survey conducted to detect microparasites it was observed that some specimens of the marine fish D. volitans showed a hypertrophy of the gallbladder with a green-brownish colour compared with the light green colour of the non parasitized gallbladders. Several developmental life cycle stages and free spores were observed immersed in the bile and located within the epithelial cells of the gallbladder wall (Figs 1-3,4-5). Based in the spore morphology, the parasite was identified as belonging to genus Myxidium Bütschli, 1882 and following Lom and Dyková (2006), we propose the establishment of a new species classified as follows: Phylum Myxozoa Grassé, 1970, Class Myxosporea Bütschli, 1881, Order Bivalvulida Shulman, 1959, Family Myxidiidae Thélohan, 1892, Genus Myxidium Bütschli, 1882. Myxidium volitans sp. nov. (Figs 1-3,4-5,6-7, 8-11, 12, 13 ) Description - Developmental stages, including immature and mature spores, with morphological characters of the genus Myxidium Bütschli, 1882 were observed free and immersed in the bile and in the epithelial cells of the gallbladder wall. The development was asynchronous with all the developmental stages and immature spores inter cellular in the epithelial cells of the gallbladder wall (Figs 4-5, 6-7 ). Some free mature spores were observed in the initial tract of the intestine. The spore bodies are fusiform, sometime slightly crescent-shaped with smooth surface and more or less rounded ends (Figs 1-3). The spore wall, is thin and smooth, comprised two equal-sized valves without ridges or projections (Figs 1-3). Sutural lines joining valves were hardly visible. Mature fresh spores have the following dimensions: 21.7 ± 0.3 µm (range 21.3-22.0) (n = 50) in length and 5.6 ± 0.4 µm (5.2-5.9) (n = 30) in width and contain two pyriform polar capsules (PC) that are 5.0 ± 0.4 (4.6-5.5) µm long and 2.3 ± 0.3 (2.0-2.5) µm (n = 30) wide, these are situated in each extremity of the spore (Figs 1-3). The PC wall measured 0.20-0.29 µm (n = 30) thick (Figs 7, 8-11). The PC wall consists of a continuous external dense layer and an internal hyaline thick material which contains an apical pore without a visible stopper (Figs 7-9). The PC contains a polar filament (PF) with irregular arrangements that is projected laterally from its apical region to the bases of PC and coiled from the bases to the tip of PC (Figs 7-11). Some regular and parallel longitudinal striations were observed at the periphery of the PF. At the matrix of the PC there are 1-2 S-like structures that are adherent to the periphery of the PF (Figs 8-11). Schematic drawings of the spore and PF based on ultrathin serial sections are shown in Figs 12, 13. Type host - D. volitans (Linnaeus, 1758) (Teleostei: Dactilopteridae) (Brazilian common name coió). Site of infection - Different developmental stages and immature spores in the epithelial cells of the gallbladder wall and later developmental stages, immature and mature spores immersed in the bile. Type locality - Atlantic coast near Niterói (22º58'S 43º00'W). Type data and depository - A glass slide with several semithin sections of the mature spores and some other developmental stages of the hapantotypes was deposited in the Myxozoa Type Slide Collection at the Instituto Nacional de Pesquisa da Amazônia - INPA, Amazonas, Manaus, Brazil, under acquisition 004/11. Prevalence of infection - Twenty two out 64 (40.7%) of D. volitans specimens [15/39 (38.5%) for females, 7/25 (28%) for males]. Pathology signs - The signs of the infection were macroscopically observed by a well evidenced gallbladder hypertrophy and by the appearance of a green-brownish colour of the infected gallbladders in contrast to the light green colour in the non-infected specimens. Some spores were observed in the initial tracts of the intestine. Etymology - The specific epithet (volitans) is derived from the name of the specific epithet of the genus of the host. DISCUSSION In the present paper we use the light microscopy and ultrastructural data to describe the spores of the myxosporean isolated from the gallbladders of the marine fish D. volitans which have widely geographic distribution in the Atlantic Ocean (Eschmeyer & Demoster 1990). The present study reveals a structural organization and similarities to genus Myxidium Thélohan, 1892 (family Myxidiidae) (Lom & Dyková 1992, 2006, MacKenzie & Kalavati 1995). This genus is characterized by having elongated spores, that are slightly crescent-shaped, with two PC in opposite ends, discharge terminally and a longitudinal curved sutural line (Lom & Dyková 2006). Considering the separation of marine and freshwater myxosporean species into two major branches (or clades) in results of the recent molecular data using small subunit DNA sequences (Kent et al. 2000, 2001, Fiala 2006), we consider for this discussion only Myxidium spp hosted in the marine Atlantic fishes. On the other hand, taking in consideration the role of tissues and organs preferences in differentiating between closely related species for identification of species (Lom & Dyková 2006), we establish spore comparison only infecting fish gallbladders from hosts collected from Atlantic areas. So, comparing the characteristics of M. volitans sp. nov. (morphology and dimensions of the spore and PCs, number of coils and arrangements of the PFs, and host species), none of these characters were simultaneously equals when compared with those reported for Myxidium spp infecting fish collected in the different geographical areas of the Atlantic Ocean (Table). The spores of M. volitans sp. nov. are substantially smaller (21.7 ± 0.3 µm long) than M. gigantissimum (97.5 µm long) and larger than M. oviform spores (11.75-16.25 µm) and than M. trachinarum spores (17.2 µm). All other spores of the different Myxidium spp reported in the Table have similar ranges and mean of dimensions, however, with different PC arrangement and polar capsule coils (PFc) number. While the spores of M. volitans sp. nov. have PF coiled 2-3 times, in all other species the PFc are coiled more times, excepted the PC of M. sphaericum that have a similar number of PFc (3-4). These two species have, however, different morphology of spores: M. volitans have fusiform slightly crescent-shaped with more or less round end, while M. sphaericum have fusiform spores with broad at the centre and a relatively blunt extremity in valvular view (MacKenzie & Kalavati 1995). Moreover, the ultrastructural analyses showed the PC and the arrangement of the PC tapering from its tip to the bases and an irregularly fold from the bases to tip of the PF, and this structural organization is different of those of the previously described species. Additionally the presence of the longitudinal striation on the periphery of the PC and the presence of the some S-like structures in the PC matrix were never reported in the previously described species. And finally, the species here described differs from the previously reported Myxidium spp (Table) in terms of its host specificity (Lom & Dyková 1992). The confusion in the literature regarding the validity and host specificity among some of these Myxidium spp recorded in the Table, have been reported some years ago. Considering the similar morphology of the spores of M. sphaericum and M. bergensis, it was suggested that two of these species may be conspecific (Noble 1957, Moser et al. 1989). Recently was referred and discussed by MacKenzie et al. (2010) the existence of a great confusion and doubt regarding the validity and host specificities of M. incurvatum, M. phaericum, M. bergensis and M. gadi which need detailed investigation on the morphological and molecular data. ACKNOWLEDGEMENTS To the technical assistance of Joana and João Carvalheiro. REFERENCES
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11095f6-7.jpg] [oc11095f1-3.jpg] [oc11095f13.jpg] [oc11095t1.jpg] [oc11095f12.jpg] [oc11095f8-11.jpg] [oc11095f4-5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}