 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 6, Sept., 2011, pp. 755-758 Short Communication Potential impact of a presumed increase in the biting activity of dengue-virus-infected Aedes aegypti (Diptera: Culicidae) females on virus transmission dynamics Paula Mendes LuzI, +; Tamara Nunes Lima-CamaraII; Rafaela Vieira BrunoII; Márcia Gonçalves de CastroIII; Marcos Henrique Ferreira SorgineIV, V; Ricardo Lourenço-de-OliveiraIII; Alexandre Afrânio PeixotoII, V ILaboratório
de Pesquisa Clínica em DST & AIDS, Instituto de Pesquisa Clínica
Evandro Chagas Received 23 March
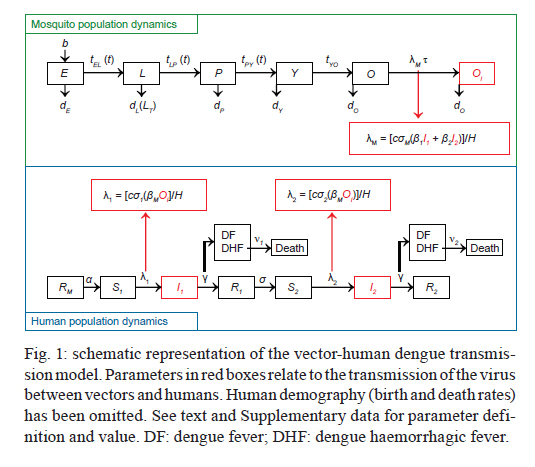
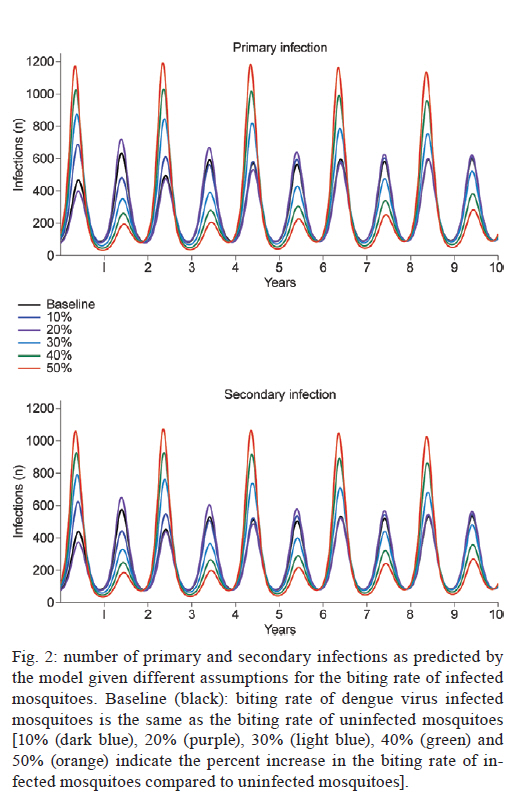
2011 Code Number: oc11127 Abstract Recently, we showed that infection with dengue virus increases the locomotor activity of Aedes aegypti females. We speculate that the observed increased locomotor activity could potentially increase the chances of finding a suitable host and, as a consequence, the relative biting rate of infected mosquitoes. We used a mathematical model to investigate the impact of the increased locomotor activity by assuming that this activity translated into an increased biting rate for infected mosquitoes. The results show that the increased biting rate resulted in dengue outbreaks with greater numbers of primary and secondary infections and with more severe biennial epidemics. Key words: Aedes aegypti, locomotor activity, dengue transmission Dengue fever is one of the most important mosquito-borne viral diseases in the world and is endemic in approximately 112 countries. It has been estimated that almost 100 million cases of dengue fever and half a million cases of dengue haemorrhagic fever occur worldwide (Gurugama et al. 2010). The main dengue vector, Aedes aegypti (Diptera: Culicidae), is a widely distributed species, occurring in many countries in the world. This mosquito is diurnal, very anthropophilic, endophilic and highly abundant in the urban districts and slums of many countries (Braks et al. 2003, Lima-Camara et al. 2006). The behaviour of insect vectors can be affected by infection with viruses and other parasites (e.g., Grimstad et al. 1980, Rowland & Lindsay 1986) and by symbionts (e.g., Evans et al. 2009, Moreira et al. 2009). There is evidence that the dengue virus can alter the behaviour of Ae. aegypti. For example, dengue-infected mosquitoes have been shown to take longer to feed than uninfected mosquitoes (Platt et al. 1997). Recently, we reported that Ae. aegypti females that were experimentally infected with dengue virus showed an increase of up to ~50% in their locomotor activity compared to uninfected controls (Lima-Camara et al. 2011). Although it is yet unknown whether this behavioural change has any effect on the biting activity in the wild, we speculate that the higher levels of locomotor activity observed in dengue-virus-infected Ae. aegypti females could potentially increase their chance of finding a suitable host and, as a consequence, their relative biting rate. In the current study, we used a simplified version of a previously developed model (Luz et al. 2009, 2011) to examine the potential impact of this increased activity level of dengue-virus-infected mosquitoes on dengue transmission dynamics. A complete description of this model is beyond the scope of this short communication and can be found in the above-mentioned references. Briefly, the model describes the vector-human transmission cycle of dengue by incorporating both mosquito and human dynamics (Fig. 1). The model's system of differential equations and the parameters are detailed in the Supplementary data. For the mosquito population dynamics, six compartments are defined. The first three mosquito life stages, egg, larva and pupa are denoted by E, L and P, respectively. The adult life stage is composed of two compartments: young mosquitoes (Y) (1-5 days of age, which do not yet lay eggs) and old mosquitoes (> 5 days of age, which lay eggs). To simulate dengue transmission, the old adult life stage is further subdivided into uninfected adults (O) and dengue-virus-infected adults (OI). The transition rates between life stages are given by tij (where i is the current life stage and j is the next life stage). Seasonality is built into the transition rates between model compartments. Weather data from Rio de Janeiro were used to parameterise the temporal pattern of the transition rates, which were mathematically defined as sinusoidal functions (Luz et al. 2009). The transition rate from egg to larva (tEL) follows the pattern for precipitation because eggs hatch in response to water. The transition rates from larva to pupa (tLP) and pupa to young adult (tPay) are functions of temperature because temperature accelerates Ae. aegypti's development rate (Scott et al. 2000b). The transition rate from young adult to old adult (tYO) is assumed to be constant. The death rate of each mosquito compartment (di) (where i = E, L, P, Y, O, OI) is the background mortality from natural sources. Field studies have shown that the larval development of Ae. aegypti is density dependent (Southwood et al. 1972, Barrera et al. 2006) and this dependence is built into the function of the background death rate of the larvae (dL). Recently, cohort studies of mosquito populations showed that adult death is age dependent (Styer et al. 2007a). Thus, for the adult life stage, the death rates vary by age group. The rate of production of viable eggs per adult is given by b (Styer et al. 2007b). O acquire infection (entering compartment OI) at a rate that depends on the force of infection (λM) and the duration of the extrinsic incubation period (τ). We defined λM as a function of (i) the biting rate (c), (ii) the proportion of infected humans [with either primary (I1) or secondary (I2) infections] and their respective probabilities of infecting a mosquito (β1 and β2) and (iii) the susceptibility of mosquitoes (σM). Thus, λM = [cσM(β 1I1 + β2I2)]/H, where H is the total number of humans. For the human population dynamics, seven compartments are defined. We incorporated the demographic dynamics of the human population from the city of Rio de Janeiro into the mathematical model. Newborns (Rm) have maternally acquired immunity that wanes at rate α. Susceptible humans (S1) acquire primary infection at the λM rate λ1, which depends on (i) c, (ii) OI and their infectivity (βM) (iii) the density of adult mosquitoes (OT) to H and (iv) the susceptibility of humans to primary infection (σ1). Thus, λ1 = [cσ1(β MOI]/H. The risk of death for individuals presenting with dengue haemorrhagic fever is given by ν1. Nonetheless, the majority of I1 recover at rate γ into a cross-immunity compartment (R1). Cross-immunity wanes at a rate δ, resulting in susceptibility to secondary infection (S2) at a rate λ2. This force of secondary infection depends on (i) c, (ii) OI and βM, (iii) OT and H and (iv) the susceptibility of humans to secondary infection (σ2). Thus, λ2 = [cσ2(β MOI)]/H. The majority of humans with I2 re-cover at rate γ into a resistant compartment (R2). Those presenting with dengue haemorrhagic fever die with probability ν2 (which is greater than ν1 because secondary infections are more severe) (Gubler 1998, Guzman & Kouri 2002). Similar to previous analyses (Bartley et al. 2002), we modelled humans as only being susceptible to two dengue infections. Despite this simplification, we incorporated the force of infection for all four serotypes by defining full susceptibility to primary infection and 75% susceptibility to secondary infection resulting from immunity to the primary serotype. For the present analysis, we assumed that the increased locomotor activity of dengue-virus-infected mosquitoes improves their likelihood of finding a suitable host and, as a consequence, enhances their biting rate, represented by parameter c described above. We simulated dengue transmission dynamics assuming a baseline value for the biting rate and assuming an increased biting rate for infected mosquitoes. The impact of the increased biting rate of infected mosquitoes was evaluated in the number of primary and secondary infections predicted by the model. Fig. 2 shows the simulation results. The increase in the activity was assumed to manifest as a percent increase of the biting rate of infected mosquitoes relative to uninfected mosquitoes. Based on our previous experimental findings (Lima-Camara et al. 2011), we explored different degrees of the relative increase in the biting rate of dengue-infected mosquitoes ranging from no increase to a 50% increase in increments of 10%. When no difference was assumed between the biting rates of uninfected and infected mosquitoes (baseline assumption, black), the model predicted annual dengue epidemics of a relatively constant size (i.e., with a relatively constant number of primary and secondary infections per year for 10 years). Overall, when assuming that the biting rate was increased, an increase in the number of infections was observed. For example, when assuming a 50% increase in the biting rate of dengue-infected mosquitoes, the percent increases in the number of primary and secondary infections were 3.8% and 6.5%, respectively, for the 10-year period shown in Fig. 2. In addition, the impact of the increased biting rate of infected mosquitoes on the transmission dynamics of dengue was profound, causing more severe biennial epidemics (colours in Fig. 2). Transmission dynamics models aim to represent, using mathematical equations, the essential components of biological processes and how these components relate to each other. The simplest model, the SIR model, represents the dynamics of a directly transmitted disease by dividing up the host population on the basis of whether they are susceptible, infectious or recovered/immune. The most important parameter of the SIR model is the rate at which susceptible hosts become infected, usually called the force of infection or the instantaneous incidence rate of infection. For vector-borne diseases such as dengue, this rate depends on parameters such as vector density, vector survival, the extrinsic incubation period of the virus and the biting rate (Nishiura 2006). In our model, these parameters are incorporated into the forces of infection acting on humans and mosquitoes. Uninfected mosquitoes acquire the infection at a rate given by the force of infection (λM) (Fig. 1). λM is a function of the biting rate (c) which is the average number of bites per mosquito per day. Similarly, acting on susceptible humans are the forces of primary and secondary infection (λ1 and λ2) which also depend on the biting rate of the mosquitoes (Fig. 1). In fact, the biting rate is the only parameter entering the formula of all three forces of infection and is thus expected to greatly impact the transmission dynamics. In our analysis, we found that the increased biting rate modified the dengue transmission dynamics simulated by the model. The absolute number of infections was greater, and most strikingly, the annual pattern of epidemics changed, with more severe biennial epidemics being observed. This pattern of biennial epidemics more closely resembles the observed multi-annual cycle of dengue epidemics in endemic countries, including the pattern observed in Brazil (Nogueira et al. 2007, Nogueira & Eppinghaus 2011). In agreement, other studies have shown the importance of the parameters related to the vector-host interaction, including the biting rate, in determining the seasonal and multi-annual cycle of dengue (Hay et al. 2000, Bartley et al. 2002). A more severe biennial epidemic implies that the virus, during the epidemic, is circulating among a greater proportion of the population, thus causing the number of secondary infections to increase. Indeed, the relative increase in the number of secondary infections was always greater than the relative increase in the number of primary infections. This finding is of clinical relevance because secondary infections have a greater likelihood of resulting in dengue haemorrhagic fever (Guzman & Kouri 2002). References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11127f2.jpg] [oc11127f1.jpg] |
| |||||||||

{kind=link}
{kind=link}