 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, No. 6, Sept., 2011, pp. 759-762 Short Communication Can thymic epithelial cells be infected by human T-lymphotropic virus type 1? Klaysa Moreira-RamosI, II, III; Flávia Madeira Monteiro de CastroI; Leandra Linhares-LacerdaI; Wilson SavinoI, + ILaboratório
de Pesquisas sobre o Timo, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365,
21040-360 Rio de Janeiro, RJ, Brasil
Received 25 April
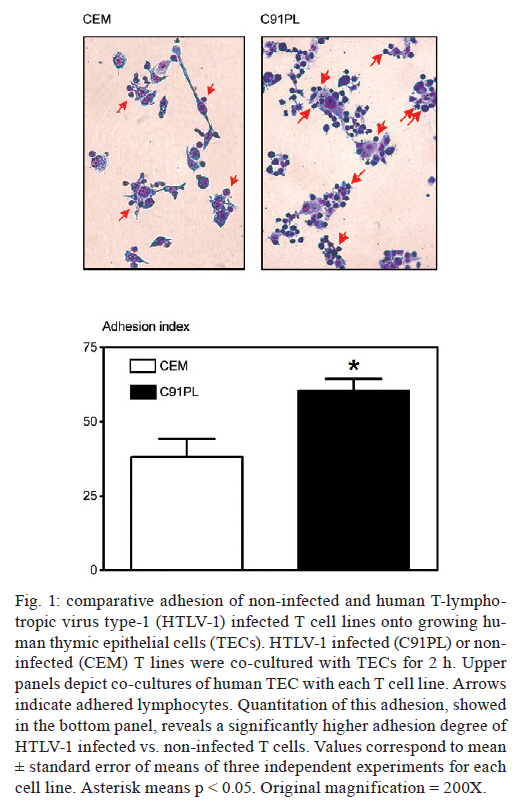
2011 The human T-lymphotropic virus type-1 (HTLV-1) is the cause of adult T cell leukaemias/lymphoma. Because thymic epithelial cells (TEC) express recently defined receptors for the virus, it seemed conceivable that these cells might be a target for HTLV-1 infection. We developed an in vitro co-culture system comprising HTLV-1+-infected T cells and human TECs. Infected T cells did adhere to TECs and, after 24 h, the viral proteins gp46 and p19 were observed in TECs. After incubating TECs with culture supernatants from HTLV-1+-infected T cells, we detected gp46 on TEC membranes and the HTLV-1 tax gene integrated in the TEC genome. In conclusion, the human thymic epithelium can be infected in vitro by HTLV-1, not only via cell-cell contact, but also via exposure to virus-containing medium. Key words: thymic epithelial cells, HTLV-1 infection, adult T cell leukaemia/lymphoma, cell adhesion The human T-lymphotropic virus type 1 (HTLV-1) infects approximately 20 million people around the world (Proietti et al. 2005), including people in Brazil. HTLV-1 infection causes various inflammatory disturbances and two different pathologies are observed: HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP), which is a neuroinflammatory disease that involves the peripheral nerves of the spinal cord (Osame et al. 1986), and adult T cell leukaemia/lymphoma (ATL), which is characterised by lymphoproliferation of mature T lymphocytes, particularly CD4+CD25+ activated T cells (Uchiyama et al. 1977). Although CD4 T lymphocytes are the main targets for HTLV-1, there is evidence that the virus can infect CD8 T lymphocytes (Nagai et al. 2001), thymocytes (Maguer-Satta et al. 1995), CD34+ progenitor cells (Feuer et al. 1996), endothelial cells (Setoyama et al. 1998), astrocytes (Mor-Vaknin et al. 1998) and dendritic cells (Jones et al. 2008). Recent studies have indicated that some receptors are required for efficient HTLV cell entrance, including glucose transporter-1 (Glut-1) (Manel et al. 2003), heparan sulfate proteoglycans (Jones et al. 2005) and neuropilin-1 (NP-1) (Ghez et al. 2006). These molecules can be found in the thymus (Murdoch et al. 1994, Swainson et al. 2005, Lepelletier et al. 2007), the primary lymphoid organ in which bone marrow-derived cell precursors undergo T lymphocyte differentiation. In particular, thymic epithelial cells (TECs), the major component of the thymic microenvironment, express large amounts of NP-1 (Lepelletier et al. 2007). Thus, it seemed conceivable that TECs might be a target for HTLV-1 infection. We approached this issue by applying an in vitro co-culture system comprising an HTLV-1+ infected T cell line, C91PL (Popovic et al. 1983); a control uninfected T cell line, CEM (Foley et al. 1965), and a foetal TEC line (Fernandez et al. 1994), which is able to adhere to developing thymocytes (Ribeiro-Carvalho et al. 2002). Cells were grown in Roswell Park Memorial Institute-1640 medium supplemented with 10% foetal calf serum, 2 mM glutamine, 100 U/mL penicillin and 100 U/mL streptomycin (Gibco, Scotland, UK) in a CO2 atmosphere at a temperature of 37ºC. Culture flasks containing 2 x 104 TECs were washed 24 h after plating and 2 x 105 cells from CEM or C91PL T cell cultures were added to the flasks and left to adhere for 2 h. Supernatants containing floating lymphocytes were discarded and the contents of each flask (TECs + adhered lymphocytes) were fixed in absolute ethanol, stained with Giemsa and counted to determine the adhesion index (AI), as previously described (Ribeiro-Carvalho et al. 2002):
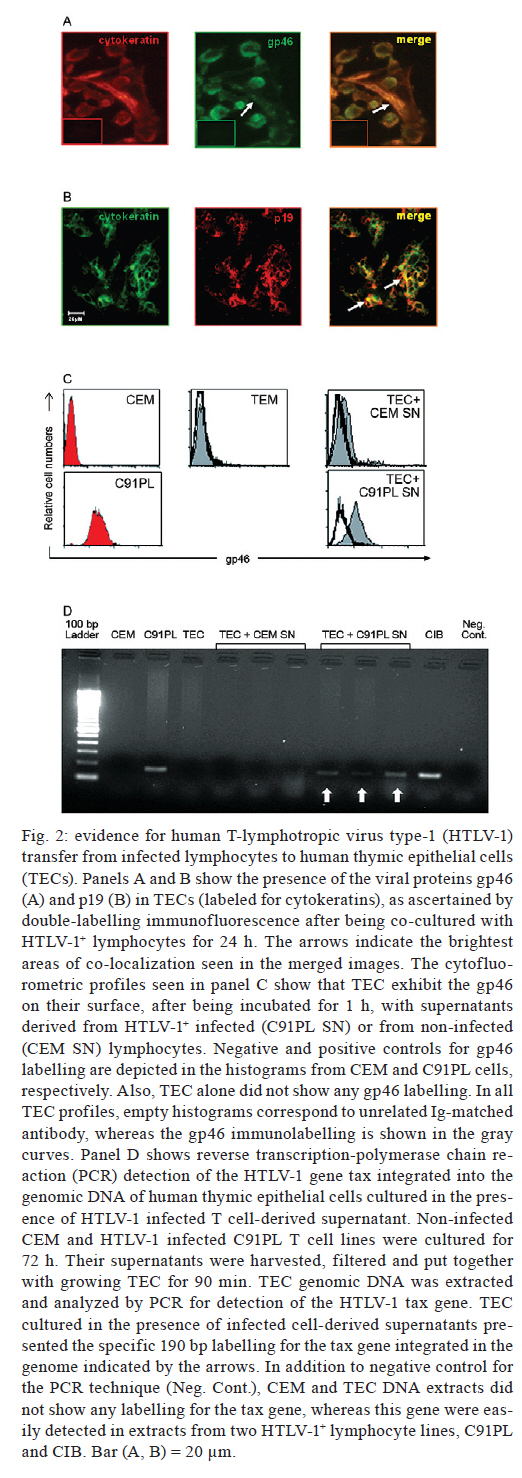
In other experiments, TECs were detached from the culture flasks using 10 mM ethylenediamine tetraacetic acid and the lymphocytes were harvested, centrifuged and submitted to cytofluorometry. For cytofluorometry analysis we used appropriate dilutions of monoclonal antibodies with the following specificities: NP-1, CCR4 (Pharmingen/Becton-Dickinson, San Diego, USA), Glut-1 (R&D Systems, Minneapolis, USA) and HTLV-1 gp46 (Abcam, Cambridge, USA). Isotype/fluorochrome-matched unrelated immunoglobulins (Becton Dickinson) were used as negative controls. Immunostaining was performed as follows: cells were incubated with phosphate buffered saline (PBS)/bovine serum albumin 0.5%/sodium azide 0.05% for 15 min, exposed to the fluorochrome-labelled antibodies for 20 min, washed in PBS and fixed in 1% formaldehyde (Ribeiro-Carvalho et al. 2002). Image acquisition and analysis were performed using a FACSCalibur® device (Becton Dickinson) equipped with CellQuest software. To determine whether viral proteins could be detected in TECs, in some experiments, TECs and C91PL cells were co-cultured for 24 h and then submitted to double-label immunohistochemistry, as described previously (Lepelletier et al. 2007). For this experiment, we used polyclonal pan-cytokeratin rabbit serum (Dako Co, Carpinteria, USA), anti-HTLV-1 gp46 and anti-HTLV-1 p19 (Abcam) monoclonal antibodies, as well as the secondary antibodies Alexa-488-labelled goat anti-mouse and Alexa-546-coupled goat anti-rabbit Ig (Amersham Biosciences, Buckinghamshire, UK). TECs were cultured for 24 h in Lab-Tek chambers and were sequentially subjected to a given primary monoclonal antibody and the corresponding fluorochrome-labelled anti-mouse second antibody, followed by incubation with the anti-cytokeratin serum, the binding of which was revealed with specific fluorochrome-coupled secondary antibodies. The slides were then analysed by fluorescence microscopy. Other experiments were designed to determine whether culture supernatants derived from HTLV-1+ T cells could convey viral elements to cultured TECs. For these experiments, lymphocytes (2 x 106/mL) were cultured for 72 h in Falcon tubes and centrifuged, and the supernatants were filtered through 0.22 µM pore size membranes, which retain cells but allow the passage of virus particles. We then plated 5 x 105 TECs in culture flasks. Twenty-four hours later, these cells were incubated for 90 min with supernatants derived from infected or uninfected lymphocytes. Cells were then harvested and labelled with antibodies against the viral proteins gp46 and p19. Alternatively, their DNA was extracted for polymerase chain reaction (PCR) analysis. For PCR, genomic DNA from TECs exposed to lymphocyte-derived culture supernatants was extracted using the Wizard SV Genomic DNA Purification System kit from Promega Co (Madison, USA), according to the manufacturer's instructions. The primers and the procedures used for PCR were derived from prior research (Lee et al. 2004). For the various experiments summarised above, quantitative data were expressed as the mean ± standard error and the results were statistically analysed using Student t test. Differences were considered statistically significant when the p values were < 0.05. In the first set of experiments, we showed that after 2 h in co-cultures, infected T cells were able to adhere to TECs, exhibiting an AI that was significantly higher than that obtained with the uninfected T cells (Fig. 1). We thus tested whether, in heterocellular cell-cell adhesion conditions, we could detect viral proteins in TECs. We co-cultured the cells for 24 h and then the TECs were stained with anti-cytokeratin and anti-gp46 antibodies. As seen in Fig. 2A, viral gp46 and cytok2ratin were co-localised in TEC cultures, indicating that these cells might have HTLV-1 on their surface or in the cytoplasm. This result was confirmed using double-label immunofluorescence for the detection of cytokeratins and the viral matrix protein p19 (Fig. 2B). We then tested the hypothesis that TECs can be infected by free HTLV-1 virions using the supernatant of infected T cells. Epithelial cells were cultured for 90 min with supernatant harvested from infected T cells cultured for 72 h. Cells were then washed, immunostained with the anti-gp46 antibody to detect this viral protein on their surface and analysed by cytofluorometry. As shown in Fig. 2C, only TECs incubated with supernatants from HTLV-1-infected lymphocytes were labelled to visualise viral gp46. Finally, we used reverse transcription-PCR to screen for the presence of the Tax gene in the genome of TECs treated with HTLV-1 infected cells. These experiments revealed that HTLV-1 was able to penetrate into TECs and integrate its proviral DNA (Fig. 2D). In conclusion, we showed herein that the human thymic epithelium can be infected in vitro by HTLV-1, not only via cell-cell contact, but also via exposure to virus-containing medium. Given that activated T lymphocytes and dendritic cells recirculate from the periphery to the thymus (Tian et al. 2007, Li et al. 2009) and that HTLV-1 can be transmitted in a cell-free manner, as has been reported for dendritic cells (Jones et al. 2008), it is conceivable that T lymphocytes and dendritic cells, once infected by HTLV-1, carry the virus into the thymus and infect microenvironmental cells, such as TECs. Thus, we hypothesize that the human thymic epithelium is a putative reservoir for HTLV-1, opening up a research field related to the role of the thymus in the HTLV-1 infection cycle. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11128f2.jpg] [oc11128f1.jpg] |
| |||||||||


{kind=link}
{kind=link}