 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 64-69 Original Article Pregnancy malaria: cryptic disease, apparent solution Patrick Emmet Duffy+; Michal Fried Laboratory of Malaria
Immunology and Vaccinology, National Institute of Allergy and Infectious Diseases,
National Institutes of Health, Rockville, MD, USA Received 2 May
2011 Code Number: oc11141 Abstract Malaria during pregnancy can be severe in non-immune women, but in areas of stable transmission, where women are semi-immune and often asymptomatic during infection, malaria is an insidious cause of disease and death for mothers and their offspring. Sequelae, such as severe anaemia and hypertension in the mother and low birth weight and infant mortality in the offspring, are often not recognised as consequences of infection. Pregnancy malaria, caused by Plasmodium falciparum, is mediated by infected erythrocytes (IEs) that bind to chondroitin sulphate A and are sequestered in the placenta. These parasites have a unique adhesion phenotype and distinct antigenicity, which indicates that novel targets may be required for development of an effective vaccine. Women become resistant to malaria as they acquire antibodies against placental IE, which leads to higher haemoglobin levels and heavier babies. Proteins exported from the placental parasites have been identified, including both variant and conserved antigens, and some of these are in preclinical development for vaccines. A vaccine that prevents P. falciparum malaria in pregnant mothers is feasible and would potentially save hundreds of thousands of lives each year. Key words:
Plasmodium falciparum - pregnancy malaria - placenta - chondroitin sulphate
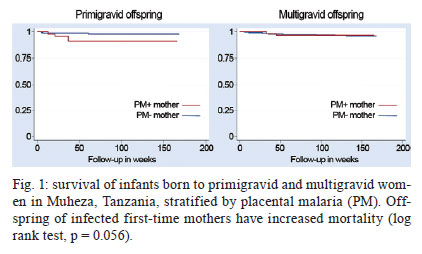
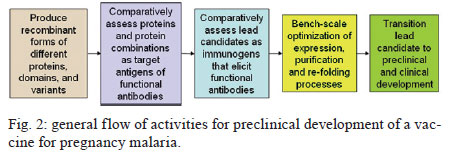
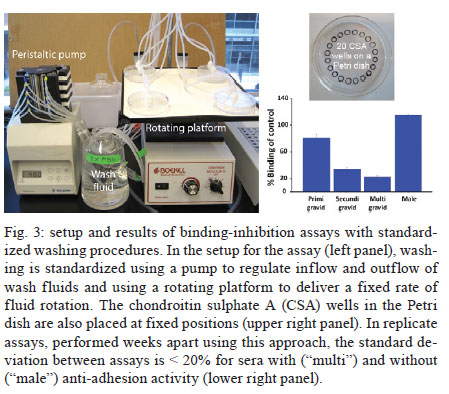
A - vaccine In this setting, a vaccine to prevent malaria during pregnancy is urgently needed. The feasibility of such a vaccine is supported by naturally acquired immunity. Women can develop immunity that prevents malaria after one or two pregnancies. This immunity corresponds to the acquisition of antibodies against placental infected erythrocytes (IE), a distinct form that adheres to the placental receptor chondroitin sulphate A (CSA) and is sequestered in the placenta. Women who acquire antibodies against CSA-binding or placental IEs have significantly improved outcomes, including increased maternal haemoglobin levels and babies with higher birth weights (Duffy & Fried 2003, Staalsoe et al. 2004). The cryptic burden of pregnancy malaria - In areas of stable malaria transmission, including much of Africa, women are semi-immune and therefore do not typically present with acute disease when infected with Plasmodium falciparum. Instead, the disease develops in the mother and offspring through an insidious process and the resulting malaria-related sequelae are problematic. The potential benefit of a vaccine to protect against malaria during pregnancy can be estimated by the impact of naturally acquired immunity and this is one approach that can be used to gain a better understanding of the burden of disease. Immune multigravid women, for example, may not receive significant benefits from malaria prevention during pregnancy because their existing immunity already prevents poor pregnancy outcomes that are caused by malaria. The known poor outcomes related to malaria include maternal anaemia, low birth weight, and perinatal and infant mortality. Recent studies of resistant multigravid and susceptible primigravid mothers indicate that pregnancy malaria causes additional problems that are not always recognised as malaria-related sequalae, including hypertension in mothers and infant morbidity and mortality that are not related to low birth weight. The benefits of a vaccine can also be estimated from studies showing the impact of antimalarial prophylaxis during pregnancy. For example, in Gambia, chemoprophylaxis with Maloprim® (pyrimethamine/dapsone) provided significant benefits, primarily to primigravid women (Greenwood et al. 1994, Menendez et al. 1994) and grandemultigravid (parity > 7) women (Greenwood et al. 1989, 1994). Compared to those receiving placebo, primigravid patients receiving Maloprim® had lower rates of parasitaemia, delivered babies with higher birth weights and had higher haematocrits. Based on its ability to increase birth weight, Maloprim® chemoprophylaxis was estimated to reduce the mortality of neonates born to primigravid women by 42% and the post-neonatal mortality of infants born to primigravid women by 18% (Greenwood et al. 1992). Prophylaxis was estimated to reduce the mortality of infants born to multigravid women by 5%, although this benefit was not significant. Neonatal and infant mortality and morbidity - A recent meta-analysis of three randomised studies showed that preventative drugs during pregnancy reduced perinatal mortality for first-time offspring (Garner & Gülmezoglu 2006). The relative risk for those receiving prevention was 0.73 (95% confidence interval 0.53-0.99), which suggests that an effective vaccine could reduce neonatal mortality by around a quarter among first-time mothers. In separate analyses, 3-8% of total infant mortality (neonatal plus post neonatal mortality), or between 100,000-250,000 infant deaths in sub-Saharan Africa, are suspected to have resulted from low birth weight associated with pregnancy malaria. Our recent studies suggest that these extrapolations may underestimate the impact of pregnancy malaria on post-neonatal infant disease and death. We conducted the first study to directly determine how PM affects post-neonatal infant mortality (Fig. 1). In Muheza, Tanzania, we recruited mother-infant pairs at delivery and followed the offspring intensively for the first years of life. Our initial analyses focused on the outcomes of single pregnancies and excluded infants with evidence of human immunodeficiency virus infection or disease. In the survival analyses, which covered up to 55 weeks of life, malaria infection during pregnancy in first-time mothers significantly increased post-neonatal mortality in their offspring (Fig. 1) (log rank test, p = 0.056 for increased mortality of infants born to infected mothers). The post-neonatal infant mortality was 9.3% for the offspring of infected first-time mothers compared to 2.6% for the offspring of uninfected first-time mothers. In this population, where 19.1% of first-time mothers had malaria at delivery, more than a quarter of the post-neonatal deaths that occurred in the infants of first-time mothers and may have been attributable to pregnancy malaria. Malaria in multigravid women did not significantly increase the mortality of their offspring (5.5% vs. 2.9%, log rank test, p = 0.77 for differences between the offspring of infected vs. uninfected multigravid women). Based on these studies, we estimate that the population attributable risk percent of post-neonatal infant mortality due to PM is 29.2% for the first pregnancy and 8.5% for all pregnancies. These calculated effects are higher than earlier estimates (earlier estimates were extrapolated from the prevalence of low birth weight). To identify additional consequences of placental malaria, we examined the effect of PM on the child's risk of developing malaria parasitaemia. Compared to other children, the offspring of infected first-time mothers had a lower risk of developing parasitaemia during infancy, while the offspring of infected multigravid women had a significantly higher risk of developing parasitaemia (Mutabingwa et al. 2005). The increased risk for offspring to develop parasitemia that is associated with pregnancy malaria is a hidden burden of the disease that was previously unrecognised that may contribute to excess infant mortality. Preeclampsia in the mother - Our studies in Muheza, Tanzania, indicate that hypertension in first-time mothers may be another previously unrecognised consequence of pregnancy malaria (Muehlenbachs et al. 2006). Among young first-time mothers (18-20 years old), malaria increased the risk of hypertension 3.1-fold (p = 0.035). Additionally, malaria-related hypertension in young first-time mothers was associated with elevated levels of the preeclampsia biomarker sFlt1, suggesting that they had preeclampsia. By comparison, older multigravid women with hypertension had normal sFlt1 levels, suggesting that they had essential hypertension. In Muheza, 19.1% of first-time mothers have malaria at delivery. This figure indicates that more than a quarter of preeclampsia cases in this population may be related to PM. Notably, the prevalence of malaria varies widely between sites and the prevalence in Muheza is relatively low compared to other sites. Therefore, the burden of malaria-related preeclampsia in other populations may be substantially higher than we observed in Muheza if the prevalence of pregnancy malaria is higher. Pregnancy malaria has been estimated to cause 10,000 maternal deaths each year due to severe maternal anaemia. Our new finding that malaria may be precipitating preeclampsia in first-time mothers suggests that malaria-related maternal mortality may be much greater than currently recognised. Therefore, additional studies are needed. An apparent solution to pregnancy malaria - Because pregnancy malaria can be a cryptic and insidious disorder, preventive tools are the optimal public health strategy. Based on our knowledge of PM immunopathogenesis, the goal for a vaccine will be to elicit functional antibodies against IEs that bind to CSA in the placenta. The rationale for this effort is that a distinct form of the parasite that accumulates in the placenta causes disease and death in mothers and infants. Mothers become resistant to pregnancy malaria as they develop antibodies against the surface proteins of this form of the parasite. A variety of proteins, including VAR2CSA and several conserved antigens, are known to be preferentially expressed by placental parasites and several VAR2CSA domains are targets of the naturally acquired antibodies that are associated with protection. These antigens may form the foundation for the development of an effective vaccine. Variant antigens expressed by pregnancy parasites - The binding of IEs to CSA and other receptors is mediated by parasite-encoded surface antigens that are variably expressed on the surface of IE. The best-known variant surface antigens are PfEMP1 proteins, a high molecular weight protein family encoded by the multicopy var gene. Each haploid parasite genome contains approximately 60 var genes. Recombinant PfEMP1 domains bind in vitro to endothelial receptors for IEs including CSA (Salanti et al. 2003, Gamain et al. 2005), implicating PfEMP1 in parasite adhesion and as a target of protective immunity. Using specific reverse transcription-polymerase chain reaction primers for each var gene, a single gene (PFL0030C) was found to be transcribed at high levels following the selection of several parasite lines for CSA adhesion (Salanti et al. 2003). The transcription of PFL0030C was higher in placental than in non-placental isolates. The alleles of PFL0030C were designated as a new var gene sub-family, var2csa (Salanti et al. 2003). VAR2CSA proteins react to sera from endemic areas in a parity-related fashion that corresponds with acquired resistance (Salanti et al. 2004). Parasites engineered with a deletion of the var2csa gene lose the ability to adhere to CSA (Viebig et al. 2005, Duffy et al. 2006). Additionally, recombinant proteins containing any of several VAR2CSA domains can bind to CSA (Gamain et al. 2005, Resende et al. 2008). Women acquire antibodies against VAR2CSA over successive pregnancies as they become resistant to pregnancy malaria (Salanti et al. 2004). VAR2CSA-specific IgG is associated with protection from one of the major adverse consequences of pregnancy malaria, which is delivery of an infant with low birth weight (Salanti et al. 2004). These data establish VAR2CSA as a leading candidate for inclusion in a pregnancy malaria vaccine. The protein encoded by var2csa is a large protein with a molecular weight of approximately 300 kD that consists of six Duffy-binding-like (DBL) domains (Salanti et al. 2003). The tertiary structure of the full-length extracellular portion of the molecule has recently been resolved and the interaction with CSA may depend on correct folding as well as interaction with of several of its domains (Srivastava et al. 2010). The recognition of conformational epitopes is therefore a likely requirement of a VAR2CSA-based vaccine that induces adhesion-blocking antibodies (Barfod et al. 2006, Bockhorst et al. 2007). With the current (and foreseeable) constraints on genetic engineering and protein production, it is not possible to manufacture whole VAR2CSA proteins. Therefore, it will be necessary to assemble smaller parts of the molecule with the desired characteristics. Although VAR2CSA is relatively conserved, it contains stretches in which amino acid sequences vary between parasite isolates and intervening stretches that are highly conserved (Dahlbäck et al. 2006). To induce antibodies that target a high proportion of placental parasites, a vaccine should target the domains of the molecule that do not vary and investigations will be needed to determine how sequence variation affects CSA and antibody binding. Var2csa sequence diversity has been analysed from 18 full-length sequences and 90 partial var2csa gene "tags" that were amplified from parasite isolates from around the world (Trimnell et al. 2006). The full-length VAR2CSA sequences have an average homology of approximately 77% at the amino acid level. VAR2CSA sequence variability can be represented by numerous polymorphic segments that are 2-15 amino acids long, assuming a limited number of different types across parasite isolates. Most of the differences between two VAR2CSA sequences are due to different polymorphic segment types along the length of the protein. The types of polymorphic segments in the parasite population are relatively stable and originated prior to the continental separation of P. falciparum isolates, leading to a globally related pool of var2csa sequence polymorphisms (Trimnell et al. 2006, Bockhorst et al. 2007). Based on combination of in silico tools, Pepscan analysis and structural modelling to dissect the antigenic properties of the CSA-binding DBL3-X domain of VAR2CSA (Dahlbäck et al. 2006), sequence variation in the DBL3-X domain of VAR2CSA primarily occurs in regions predicted to be flexible loops under strong positive selection. The epitopes of human IgG are also predicted to be in the flexible loops, which are accessible to antibodies on the IE surface. These results and comparative sequence analyses are consistent with a model of evolution in which new var2csa sequence polymorphisms are generated by mutation and "shuffling" of polymorphic segments between var2csa sequences, thereby creating combinatorial diversity at surface-exposed loops. Conserved proteins of placental parasites - In addition to the variant antigen VAR2CSA, eight other conserved genes are also upregulated in maternal parasites (Francis et al. 2007, Fried et al. 2007). All but one of these genes encodes proteins with a putative export sequence and/or transmembrane (TM) domain and all of them are highly conserved with no known function. In previous studies, antisera to proteins encoded by two of these genes (PFI1785w and PFB0115w) did not react with the surface of maternal iRBC (Fried et al. 2007). However, our on-going studies suggest that one or more of these proteins may be exported to the surface of IEs by the parasites that infect pregnant women and that women may acquire antibodies against these proteins over successive pregnancies. In addition to PFI1785w and PFB0115w, these genes include PFB0280w, PFD1140w and PF10_0013. PFB0280w is predicted to be a 302 kD protein that will therefore need to be expressed as smaller individual domains to develop immunogens. In silico modelling using TMpred suggests that this protein contains two TM domains that flank a short domain predicted to be internal and two approximately 120 kD domains that are potentially expressed on the surface. PFD1140w is predicted to be a 40.8 kD protein and to contain a Pexel sequence, a signal peptide sequence and one TM domain at the N-terminus. The preferred model of this protein contains one TM domain with the N-terminus facing the cytoplasm and surface expression of the remaining protein. PF10_0013 is predicted to be a 27 kD protein, a size typically amenable to recombinant expression. The predicted protein contains a Pexel-like motif downstream of its signal peptide and is encoded by a two-exon gene. The leucine at the third residue of the Pexel motif has been replaced by an isoleucine in PF10_0013. If this protein is cleaved at its Pexel sequence, as occurs with other Pexel -containing proteins translocated across the vacuolar membrane, its size is predicted to be approximately 22 kD. Interestingly, a protein of approximately 22 kDa was previously detected on the surface of CSA-binding iRBC using photoactivatable radiolabelled CSA (Gowda et al. 2007). Of note, PF10_0013 lacks a predicted TM domain or GPI anchor sequence; therefore, PF10_0013 might need to interact with a membrane protein (such as VAR2CSA) for surface expression. Each of these proteins is encoded by a single copy gene and the sequence of each is completely conserved between different parasite isolates. These are attractive features for a vaccine candidate. Translating antigens into vaccines for pregnancy malaria - Our goal is to develop recombinant proteins that elicit functional antibodies against placental parasites and to transition these into a vaccine that will prevent malaria and improve pregnancy outcomes. The rationale for this effort is that a distinct form of the parasite infects the placenta and causes disease and death in mothers and infants. Mothers become resistant to pregnancy malaria as they develop antibodies against the surface proteins of this form of the parasite. A variety of proteins, including VAR2CSA and several conserved antigens that are preferentially expressed by placental parasites, have been identified and some of these, including VAR2CSA domains, are targets of the naturally acquired antibodies that are associated with protection. Preclinical development of a vaccine for pregnancy malaria is currently focused on recombinant forms of VAR2CSA and the conserved proteins expressed by placental parasites, and on determining which protein or combination of proteins are the best immunogens for eliciting functional antibodies against placental parasites (Fig. 2). The key questions for us and other scientists working on a vaccine for pregnancy malaria include: (i) which proteins, including which VAR2CSA domains and variants, will best elicit functional antibodies?; (ii) which combination of proteins, domains and/or variants will be needed for broad reactivity?; (iii) which expression system produces immunogen that elicits broadly reactive functional antibodies? The optimal vaccine construct should induce long-lasting or rapidly-boosted antibodies that abolish the binding of all placental parasites. Such a construct might be designed with sequences from VAR2CSA or other conserved pregnancy malaria proteins that will elicit broadly reactive protective antibodies, which may require correctly folded constructs. For VAR2CSA, this will require the production of sub-unit constructs that accurately mimic in vivo molecular conformations because technical constraints prevent the industrial manufacture of the entire high-molecular weight antigens. Insight into the structure and variability of the native protein can assist this process and the recent determination of the VAR2CSA structure using the full-length extracellular domain is now being used to enhance the design of immunogens based on this molecule. Assay development: monitoring vaccine success in the laboratory - Our primary assay to assess antisera is the binding-inhibition assay, which measures anti-adhesion antibodies (antibodies that block IE adhesion). Naturally acquired anti-adhesion antibodies in mothers correlate with resistance to placental infection and to an increased birth weight. Immunogens can be assessed according to their ability to elicit broadly reactive anti-adhesion antibodies in animals. A standardised binding-inhibition assay, including negative and positive control sera, is feasible in most laboratories using CSA-selected isolates of P. falciparum. This assay allows for the direct comparison of antisera raised against different immunogens for the ability to block adhesion of a panel of CSA-binding laboratory isolates. Leading candidate molecules that emerge from these studies can be assessed directly against fresh placental IEs at research facilities in endemic areas. The binding-inhibition assay has become a standard tool in the field of pregnancy malaria since we first developed it (Fried et al. 1998). The binding-inhibition assay can measure the ability of antibodies or antisera to inhibit the binding of IE to CSA or to placental sections. We have been optimising several assay parameters during the process of developing a standardised assay. As in the original technique, purified CSA is immobilised by adsorption on Petri dishes in 30 µL spots to form numerous "wells" that support IE binding. For earlier studies, IEs were prepared as 2-50% parasitaemia and 1-6% haematocrit for use in these assays. For more recent studies of antisera raised against candidate vaccine immunogens, IEs are prepared as 20% parasitaemia and 0.1% haematocrit. IE are pre-incubated with immune sera at a 1:5 dilution and then allowed to settle on the individual CSA "wells" for 30 min. Next, the sample is washed to remove non-adherent cells and the bound IEs are counted. The number of IEs bound in the presence of test sera is compared to the number bound in the presence of control non-immune serum. If the test sera have anti-adhesion activity, then the number of IE bound in the presence of test sera will be significantly lower than the number bound in the presence of control sera. A major source of variance within and between assays is due to variable washing of the adherent cells. We have focused on optimising and standardising this process along with other components of the assay. The parasites are grown to 4-8% parasitaemia in 4% haematocrit. When parasites reach the trophozoite stage, they are ready for use. The CSA "wells" are placed on the Petri dish using a template, so that each well is equidistant from the outer edge of the dish (Fig. 3). The adsorption of CSA occurs in a chamber overnight at 4ºC and is performed with 100% humidity during all steps to prevent unequal evaporation effects. The parasites pre-incubated with test or control sera are allowed to settle on the individual CSA wells (blocked with bovine serum albumin) for no more than 15 min at 37ºC in the humidity chamber. The binding-inhibition plates are then washed using a rotating platform and a peristaltic pump to regulate the washing rotation and the inflow/outflow of wash solution, respectively. The peristaltic pump delivers phosphate buffered saline (PBS) at a fixed rate through the inlet tubing without disturbing settled layers of parasitized red cells. When the level of the PBS reaches the outlet tubing, the PBS is removed from the plate at the same rate that it is being added. The rotating platform is switched on at a fixed rate, which ensures that all wells are washed with equal force. After a standard period of washing, the plates are removed from the platform and all PBS is removed. The adherent cells are fixed, dried, stained and analysed by microscopy using custom-programmed automated counting software. A vaccine that prevents P. falciparum malaria in pregnant mothers is feasible and would save hundreds of thousands of lives each year if delivered to women before their first pregnancy. Existing malaria vaccines will not protect pregnant women and their unborn children. Pregnancy malaria is caused by P. falciparum-IEs that bind to the placental receptor CSA; these IEs are then sequestered in the placenta where they cause disease and death for the mother and her offspring. Women become resistant to pregnancy malaria as they acquire antibodies that target surface proteins of placental parasites. Surface proteins of placental IE have been identified and are targets of the functional antibodies acquired by immune multigravid women. Our understanding of PM pathogenesis and immunology provides a context to guide the development of vaccines for pregnancy malaria and, more importantly, informs functional assays to assess their potential efficacy. Our knowledge, tools and targets make it possible and necessary to solve the longstanding problem of pregnancy malaria due to P. falciparum. Acknowledgements To Dr Andrew Oleinikov, for performing protein modelling using TMpred, to Melissa Bolla, for analysing the relationship of pregnancy malaria to the survival of infant offspring, to Richa Chaturvedi, Amber Randall and Joe Lograsso, for optimising the binding-inhibition assay, and to the clinical team in Muheza, Tanzania, for caring for the mothers and infants and for management of the samples. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11141f2.jpg] [oc11141f1.jpg] [oc11141f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}