 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 70-78 Original Article Biomarkers for susceptibility to infection and disease severity in human malaria Bruno Bezerril AndradeI; Manoel Barral-NettoII, III, IV, + ILaboratory
of Parasitic Diseases, National Institute of Allergy and Infectious Diseases,
National Institutes of Health, Bethesda, MD, USA Key words:
malaria - Plasmodium - biomarker - inflammation
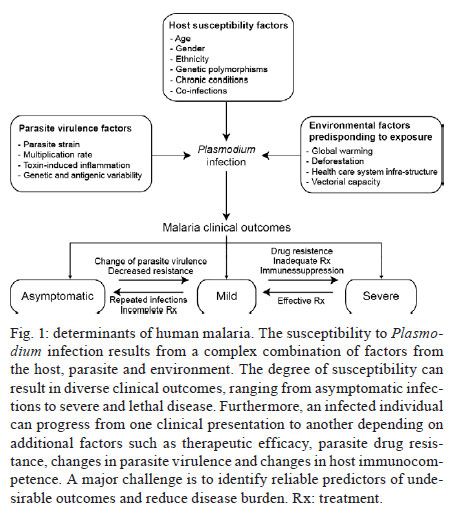
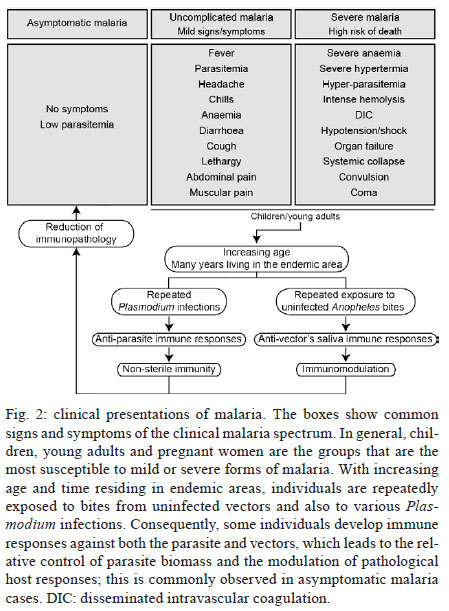
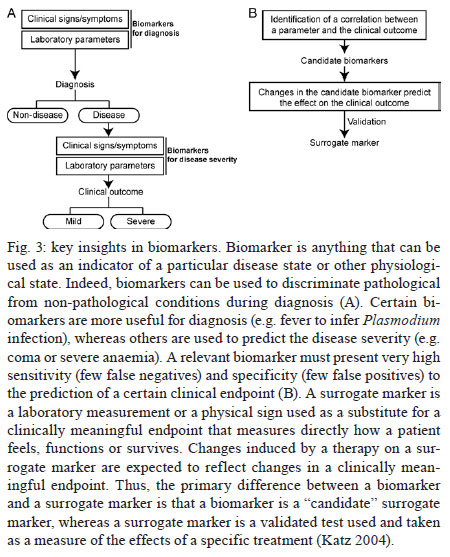
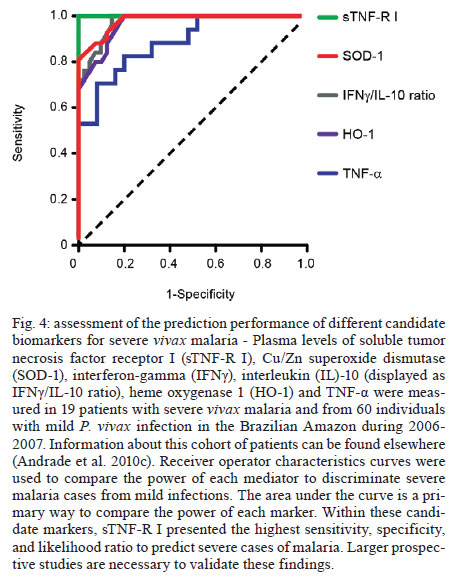
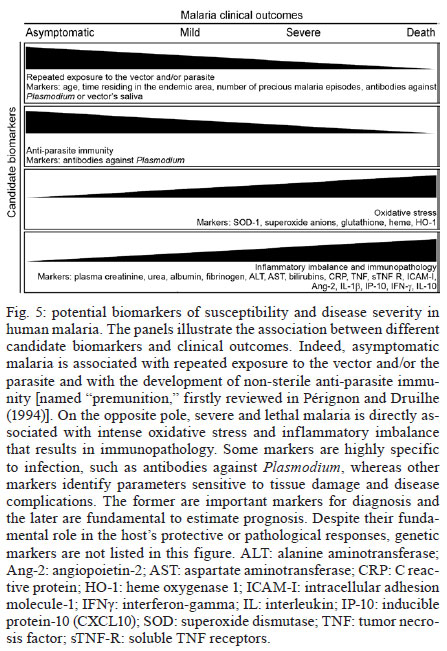
Once infected with Plasmodium, a human host generally presents with one of three major clinical outcomes: asymptomatic infection, mild disease or severe disease. These diverse clinical presentations are determined by complex interactions between the host, the parasite and environmental factors (Fig. 1). Parasite determinants include the different pathogenic potential of the various Plasmodium species. For example, P. falciparum is well known to cause severe infections more frequently than other Plasmodium species because P. falciparum exhibits a number of unique characteristics that favour increased disease severity, including high multiplication rates in both erythrocytes and reticulocytes, strong cytoadherence to infected erythrocytes [reviewed in Schofield (2007)] and toxin-induced activation of inflammatory responses [reviewed in Clark and Cowden (1999)]. Interestingly, cases of severe malaria caused by P. vivax (Andrade et al. 2010c) or uncommon Plasmodium species (Cox-Singh et al. 2010) display patterns of inflammation and immunopathology similar to those seen in severe falciparum malaria cases. These findings suggest that different Plasmodium species can trigger strikingly similar host responses that can result in severe disease. A number of host factors have already been described as important determinants of clinical outcomes in malaria cases, including age and gender, ethnicity, concomitant chronic conditions, co-infections and diverse genetic polymorphisms. Environmental factors include deforestation, health care system infrastructure and access to effective antimalarial treatment, vector exposure and sociocultural factors. Within the clinical spectrum of Plasmodium infections, we consider the two opposite poles that remain to be fully understood of utmost importance: asymptomatic malaria, which is directly associated with clinical immunity to infection and severe malaria, which underlies key processes in susceptibility to infection. Several epidemiological studies have shown that cases of asymptomatic malaria are highly prevalent in many endemic regions (Cucunubá et al. 2008, Baliraine et al. 2009, Marangi et al. 2009, Steenkeste et al. 2010). Individuals with asymptomatic Plasmodium infection can exhibit low parasitaemia for up to 60 days (Alves et al. 2002). Because of the absence of symptoms, these infected individuals do not seek health care at malaria reference centres. Furthermore, active detection of asymptomatic malaria cases is hampered by the low sensitivity of microscopy-based detection techniques in identifying very low parasite burdens (Andrade et al. 2010b). Moreover, despite low concentrations of circulating parasites, individuals with asymptomatic malaria infections can transmit Plasmodium to uninfected vectors (Alves et al. 2005). This phenomenon suggests that individuals with asymptomatic Plasmodium infections may serve as an important parasite reservoir in endemic areas. Major factors associated with asymptomatic malaria are related to the magnitude of parasite and vector exposure (Bejon et al. 2009). Indeed, it is well established that the occurrence of asymptomatic malaria is associated with increased age (Baird et al. 1991), time of residence in the endemic area and number of previous malaria episodes (Rogier & Trape 1995). After many years of repeated infections, the host develops clinical immunity against Plasmodium. In these cases, the onset of symptoms is prevented by limiting parasite burden and controlling inflammation (Fig. 2). Intriguingly, such asymptomatic carriers have developed just enough immunity to protect them from malarial illness, but not from malarial infection. Thus, the search for biomarkers of clinical immunity must explore tools to estimate both parasite/vector exposure and the host's own protective responses. Important general concepts related to biomarkers are presented in Fig. 3. The hallmark symptom of malaria is fever, which can be followed by a wide range of other symptoms including headache, chills, diarrhoea, lethargy, coughing fits and abdominal or muscular pain (Greenwood et al. 2005). Further, more severe manifestations of malaria also vary and include anaemia, hypoglycaemia, hypotension, intense haemolysis, metabolic acidosis, spontaneous bleeding, hepatitis, acute kidney failure, respiratory distress, convulsion, coma and multiple organ failure (WHO 2000). Notably, while the parameters defining severe malaria are standardised with regard to P. falciparum infections, there are as yet no consensus criteria measuring severity of vivax malaria infections. One simple approach that has been explored is based on outcomes of clinical and laboratory tests, which are used to predict overall mortality and specific complications (Mishra et al. 2007, Winkler et al. 2008). Unlike in cases of asymptomatic malaria, parasitological diagnosis is not a major concern in severe cases, as most patients present with high levels of parasitaemia that are easily detected by microscopy-based techniques. In these situations, the critical point is to exclude mixed Plasmodium infections and other comorbidities, such as bacterial sepsis, leptospirosis, dengue, viral hepatitis and acquired immune deficiency syndrome, that can result in similar clinical presentations. Thus, the search for highly specific markers should be prioritised to facilitate diagnosis. Nevertheless, less specific markers, such as those that indicate inflammatory status, are also extremely important in the light of their high sensitivity in predicting disease severity (Andrade et al. 2010d). Intense research focused on the immunopathogenesis of severe malaria will contribute to the identification of useful surrogate and specific biomarkers for cases of severe malaria. The identification of reliable biomarkers for susceptibility to infection and for disease severity can provide important insights into the diagnosis and management of malaria. Here, we review recent research advances in the identification of malaria biomarkers, focusing on biomarkers specifically related to parasite exposure and susceptibility to disease. We also present a critical analysis of the priorities for research regarding biomarkers for severe malaria. Biomarkers of exposure - Plasmodium is transmitted through the bites of Anopheles mosquitoes. As the insect takes a blood meal, arthropod saliva is injected into the vertebrate host. Vector saliva is essential to the Plasmodium life cycle (Choumet et al. 2007), facilitating blood feeding by inhibiting host coagulation and inflammatory responses [reviewed in Andrade et al. (2005)]. These effects may be directly linked to changes in host immune response (Depinay et al. 2006); furthermore, several of the active molecules present in vector saliva are immunogenic to vertebrate hosts, resulting in the initiation of an anti-saliva immune response. Immediate, delayed and systemic hypersensitivity reactions to vector saliva have been described [reviewed by Andrade et al. (2005)]. Vertebrate responses to arthropod salivary components have been used as epidemiological markers for vector exposure for ticks (Schwartz et al. 1991), phlebotomines (Barral et al. 2000, Gomes et al. 2002), Triatoma (Nascimento et al. 2001), Glossina (Poinsignon et al. 2008b) and Aedes mosquitoes (Remoue et al. 2007). In the case of malaria, the host immune response against vector saliva can be used as a biomarker of exposure to Anopheles. Two points related to the detection of biomarkers in saliva must, however, be clarified. First, detection of a host immune response against saliva does not necessarily imply host protection. This is a controversial point, as both host-protective (Donovan et al. 2007) and non-protective (Kebaier et al. 2010) findings have been reported. Currently, we utilise anti-Anopheles saliva immune responses as a marker of exposure of potential epidemiological significant. Second, the evaluation of host responses against mosquito saliva should not be confounded with diagnostic approaches that detect plasmodial products in host saliva (Wilson et al. 2008, Nwakanma et al. 2009, A-Elgayoum et al. 2010, Buppan et al. 2010, Gbotosho et al. 2010). Antibodies against Anopheles gambiae saliva have been described in young children from a region in Senegal with seasonal malaria transmission (Remoue et al. 2006). Travellers transiently exposed to An. gambiae bites in endemic areas of Africa also develop anti-saliva antibodies (Orlandi-Pradines et al. 2007). Outside of Africa, anti-Anopheles dirus salivary protein antibodies occur predominantly in patients with acute P. falciparum or P. vivax malaria; people from non-endemic areas do not carry such antibodies (Waitayakul et al. 2006). In the Americas, the presence of anti-Anopheles saliva antibodies has been described in malaria-endemic areas. Adult volunteers from communities in the state of Rondônia, Brazil, were tested for antibodies against Anopheles darlingi salivary gland sonicates (SGS), as this species is a prevalent malaria vector in Brazil. Individuals infected with P. vivax presented higher levels of anti-SGS antibodies than did non-infected individuals. This test is potentially useful as an epidemiological tool; antibody levels could discriminate between infection and non-infection with a high likelihood ratio (Andrade et al. 2009). Using a similar approach, a recent study associated the presence of anti-Anopheles albimanus saliva antibodies with mosquito bite exposure in Haiti, but did not connect these results with clinical outcomes (Londono-Renteria et al. 2010). The practical large-scale application of serological tests to detect vector saliva-specific antibodies for epidemiological purposes has been hampered by the difficulty in obtaining sufficient quantities of vector saliva and by problems in the standardisation of the reagent. Recently, the feasibility of using peptide markers to identify exposure to Anopheles bites has been demonstrated (Poinsignon et al. 2008a). Biomarkers of susceptibility - There is now strong evidence that high frequencies of genetic disorders, predominately those involving haemoglobin and red blood cell metabolic pathways and membranes, reflect relative resistance to malaria. One of the most famous genetic disorders associated with protection against death or severe disease is the sickle cell trait, haemoglobin S (HbS). The gene for HbS is widely distributed throughout many tropical regions, including sub-Saharan Africa, the Middle East and India. The first study to propose the relationship between the HbS variant and falciparum malaria showed high frequencies of the sickle cell trait in non-infected children from highly endemic areas and a relative rarity of HbS in individuals with severe disease [reviewed in Weatherall (2008)]. Although the mechanism of protection remains unknown, it has been established that the parasitised erythrocytes from a patient with sickle cell trait are much more susceptible to sickling compared to non-parasitised cells (Luzzatto et al. 1970). Thus, the removal of sickling red cells in the spleen increases the rate of parasite clearance, reducing parasitaemia and therefore the magnitude of symptoms. In addition, sickle cell carriers had significantly higher titres of IgG antibodies against a panel of malaria antigens compared to non-carrier children (Cabrera et al. 2005). A range of other possible mechanisms, however, may protect sickle cell trait carriers from severe malaria and these should be explored in further studies. Many other red blood cell mutations, such as haemoglobin C (Fairhurst et al. 2005) and alpha thalassaemia (Allen et al. 1997, Mockenhaupt et al. 2004, Williams et al. 2005) are associated with some degree of protection against falciparum malaria. The mechanisms of protection against malaria are commonly linked to reduced parasite replication within infected cells or the enhanced clearance of infected erythrocytes resulting from abnormal erythrocyte fragility (Pasvol et al. 1978, 1982, López et al. 2010). For other genetic disorders, the mechanism of protection is less well understood; this is the case for glucose-6-phosphate-dehydrogenase (G6PD) deficiency, the most common enzymopathy worldwide (Nkhoma et al. 2009). It is likely that this condition enhances red blood cell fragility as a result of increased susceptibility to oxidative stress (Cappadoro et al. 1998). Although it is protective against malaria, G6PD deficiency is also linked to a severe haemolytic syndrome triggered by primaquine, a primary drug used to treat P. vivax infections (Cappadoro et al. 1998). Beyond advancing the understanding of protective mechanisms, more effort is needed in applying this knowledge to the development of practical prophylactic or therapeutic tools. The Duffy blood group antigen illustrates how genetic epidemiology can drive essential insights into the molecular basis of malaria. Epidemiological studies first suggested that individuals lacking the Duffy antigen/chemokine receptor (DARC) were relatively protected against vivax malaria (Miller et al. 1976). Later evidence revealed that P. vivax molecules bind to DARC and are important for parasite invasion (Chitnis & Miller 1994). Recent studies have shown that P. vivax infections are common in Duffy-negative people (Cavasini et al. 2007a, b, Ménard et al. 2010), bringing attention to the question of the real role of Duffy antigens in Plasmodium infectivity. In addition to genetic determinants, there are a number of other interesting markers of malaria susceptibility. One such marker is the initial inflammatory response to Anopheles bites. Levels of antibodies against An. gambiae saliva were higher in young children who developed clinical malaria episodes than those that remained asymptomatic; these findings were interpreted to mean that the anti-saliva response is a potential marker for the risk of falciparum malaria (Remoue et al. 2006). Additionally, individuals with asymptomatic P. vivax parasitaemia presented higher anti-saliva antibody titres and a lower interferon-gamma (IFNγ)/interleukin (IL)-10 ratio than symptomatic individuals; furthermore, the IFNγ/IL-10 ratio was inversely correlated to anti-SGS titres in asymptomatic individuals (Andrade et al. 2009). Unfortunately, there is little data regarding cellular immune responses against salivary antigens; such responses are commonly observed in other diseases such as leishmaniasis (Gomes et al. 2002). It is important to verify whether the manipulation of a host's immune response against vector saliva can influence susceptibility to infection. It is well established that antibodies and T cells have crucial roles in protective immunity against the different life cycle stages of Plasmodium (Good et al. 1998). Antibodies against merozoite surface molecules inhibit parasite invasion of new erythrocytes (Giha et al. 2000). Some classes of antibodies, mainly the IgG isotype, directed against antigens of P. falciparum erythrocytes are important in antimalarial immunity; the transfer of serum from an immune person to a non-immune is protective against infection (McGregor 1964). Humoral responses can also target sporozoites, reducing parasite invasion of hepatocytes (Hisaeda et al. 2005) or, as the case of anti-P. falciparum merozoite surface protein 1, humoral responses can interfere with the attachment of infected erythrocytes to the endothelium (Giha et al. 2000). Furthermore, specific antibodies against glycosylphosphatidylinositol, an immunostimulatory molecule, suppress macrophage activation, thus decreasing cytokine production and inflammation (Schofield et al. 2002). Additionally, host antibodies directed against gametocytes interfere with the transmission of parasites to new vector mosquitoes. Although this type of immunity does not protect infected individuals, it may help to reduce infection at the community level. Biomarkers of severity - Severe malaria is a highly lethal condition that requires intensive care. As a result of their immunocompromised state, children and pregnant women are the most susceptible to complications from P. falciparum infection. Although severe cases of vivax malaria are also associated with younger age (Andrade et al. 2010c), adults appear to be susceptible even in regions with moderate to high endemicities. Multiple factors have been implicated in the pathogenesis of severe malaria caused by either P. vivax or P. falciparum infection, including uncontrolled cytokine production (Clark et al. 2006), intense haemolysis (Yeo et al. 2009, Andrade et al. 2010a) and erythropoiesis suppression (Lamikanra et al. 2007). In addition, systemic endothelial damage and obstruction of blood flow are important contributors to lethal outcomes, mainly in P. falciparum infections (Schofield & Grau 2005, Schofield 2007). Several different biomarkers have been described as useful predictors of severe malaria and many of these were discovered based on understanding of Plasmodium immunopathogenesis. The majority of what is known concerning severe malaria is related to P. falciparum infections, most likely because of this parasite's large disease burden in Africa. Because P. vivax infection has been relatively neglected compared to falciparum malaria, the factors involved in its pathogenesis are less well understood; furthermore, biomarkers for vivax malaria susceptibility and disease severity are largely unknown. Many factors have been associated with severe forms of malaria and most efforts have focused on distinguishing cerebral malaria (CM) and severe malarial anaemia from other diseases caused by uncomplicated infections. One simple factor that can influence malaria severity is parasitaemia. In P. falciparum infections, parasitaemia is directly linked to the severity of symptoms (WHO 2000). In vivax malaria, the limit of parasitaemia that can trigger pathological events is lower, so patients with relatively few circulating parasites can exhibit the inflammatory cytokine overproduction that results in fever and other complications (Karunaweera et al. 1992, Hemmer et al. 2006). In addition, as P. vivax preferentially infects reticulocytes, parasitaemia rarely exceeds 2% of circulating red blood cells and high parasite burdens are not a common feature of severe vivax malaria. Indeed, in two studies carried out in the Brazilian Amazon, patients with severe vivax malaria presented diverse parasite burdens (Alexandre et al. 2010, Andrade et al. 2010c). Differentiating CM from other causes of serious illness in endemic areas is complicated by the non-specific clinical presentation and the high prevalence of incidental P. falciparum parasitaemia (Conroy et al. 2010). Knowledge of the pathogenesis of CM and the identification of cytoadherence in P. falciparum and P. vivax (Carvalho et al. 2010) led us to test mediators associated with endothelial activation or damage. The angiogenic factor angiopoietin-2 is released from endothelial cells with reduced nitric oxide bioavailability in cases of CM (Lovegrove et al. 2009); it contributes to endothelial activation, sequestration of parasite biomass, impaired perfusion and poor clinical outcome (Yeo et al. 2008). Other strong predictors of CM that can be measured in plasma are intracellular adhesion molecule-1 (Conroy et al. 2010), interferon gamma-inducible protein 10 (IP-10), soluble tumour necrosis factor receptors (sTNF-R) and soluble FAS molecules. These molecules were independently associated with increased risk of CM-associated mortality (Jain et al. 2008). Many other markers, including IFNγ, IP-10, IL-8, MIP-1β, IL-1RA, FAS-L, sTNF-R1 and sTNF-R2, could be identified in the cerebrospinal fluid of patients with lethal CM, suggesting their involvement in CM neuropathology (Yeo et al. 2008). Recently, cell-specific microparticles, which indicate cellular damage, were demonstrated to be biological markers for cerebral dysfunction in human CM (Pankoui Mfonkeu et al. 2010). Interestingly, elevated plasma concentrations of microparticles were also found in individuals with non-complicated P. vivax infection in a study conducted in the Brazilian Amazon (Campos et al. 2010), suggesting that platelet-derived microparticles may play a role in the onset of acute inflammatory symptoms of vivax malaria. These results also suggest that the microparticles are a potential biomarker for severity of malaria caused by P. falciparum or P. vivax. Many of these candidate biomarkers could be combined in a multiplex immunoassay, which could be validated in broader applications. IL-10, together with TNF-α and erythropoietin, has been described as a good predictor of severe anaemia (Ageely et al. 2008, Ouma et al. 2008). Recently, the IFNγ/IL-10 ratio has been successfully used as a marker for pathological inflammatory activity in patients with varying disease severity (Andrade et al. 2010c). The most famous inflammatory marker of severe malaria is TNF-α, which is tightly associated with fever, paroxysms, anaemia, CM (Karunaweera et al. 2003, Armah et al. 2005) and many other systemic symptoms of the infection. It is worthy of note that as all of these markers are based on immunopathogenesis and not on the presence of the parasite itself, they have limitations in their use. After differential diagnosis, but before the onset of severe symptoms, some biomarkers can be used as predictors of complications or mortality, which could prove important in the management of potentially lethal infections. Accordingly, factors that indicate not only inflammatory status but also oxidative stress and damage to specific organs can easily be used in predictive models; they may serve as surrogate markers of disease severity. In addition, as the objective of the prediction is to influence the clinical outcome, the specificity of these markers is not critical. Indeed, non-parasite-specific markers have been shown to be powerful tools. Haemolysis, for example, is inherent to the Plasmodium life cycle during the symptomatic phase of the disease (Ekvall et al. 2001). The acute haemolysis observed in severe malaria cases results in abnormally high concentrations of free heme, which subsequently induces the production of superoxide radicals (Delmas-Beauvieux et al. 1995), cell death (Gozzelino & Soares 2011) and an imbalance between pro-inflammatory and anti-inflammatory mediators (Andrade et al. 2010a). Recently, we have shown that haemolysis related to high plasma levels of indirect bilirubin, total heme and Cu/Zn superoxide dismutase (SOD1) is consistently associated with severe vivax malaria (Andrade et al. 2010a). Furthermore, SOD1 is a better predictor of severe vivax malaria than are plasma levels of TNF-α (Andrade et al. 2010d), but this biomarker cannot be used to discriminate between P. vivax and P. falciparum infections. In a recent study of a cohort of patients with different clinical presentations of vivax malaria, we described a series of promising markers of severe disease (Andrade et al. 2010c). These markers can be easily measured and can be used to generate a score that is strongly associated with unfavourable outcomes. The markers included plasma creatinine, urea, C reactive protein, fibrinogen, bilirubins, hepatic transaminases and heme oxygenase-1. Some of these mediators were evaluated and found to have very high discriminating power (Fig. 4). A summary of some promising biomarkers of clinical protection or disease severity is illustrated in Fig. 5. Future perspectives - Although much published data exists regarding new biomarkers for malaria susceptibility and disease severity, few studies provide validation of their results using large samples or multi-centre approaches. Coordinated initiatives are necessary to design robust validation studies and generate standardised guidelines. One potential approach is the screening for susceptibility signatures using high-throughput analyses. Another method is the creation of tools using combinations of powerful biomarkers to quantify the degree of malaria susceptibility and severity. The use of a combination of biomarkers instead of one specific candidate could increase reliability and predictive power. Network analyses are necessary to address this demand, as they can provide an understanding of the real relationship between candidate biomarkers and pathological processes or clinical and parasitic determinants. Furthermore, we should emphasise larger epidemiological human studies that examine the vector-host-parasite relationship to reveal potential markers. Caution, however, is needed in the search for the ideal biomarker of malaria severity. Validation studies must be designed to discriminate between severe cases caused by P. vivax and P. falciparum. Until now, the research on falciparum malaria has been much more advanced, mostly because of the higher incidence of lethal cases. Consensus criteria to classify severe vivax malaria are now necessary; lacking such criteria, candidate biomarkers validated in studies with falciparum malaria may show weak or no correlation with vivax malaria severity and vice versa. Aside from investigating genetic determinants of the disease, the search for good predictors of susceptibility or severity must involve the generation of simple and cheap tools that can be implemented in undeveloped endemic areas. Acknowledgements To Dr Daniel L Barber, for critical discussions. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11142f5.jpg] [oc11142f4.jpg] [oc11142f3.jpg] [oc11142f1.jpg] [oc11142f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}