 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
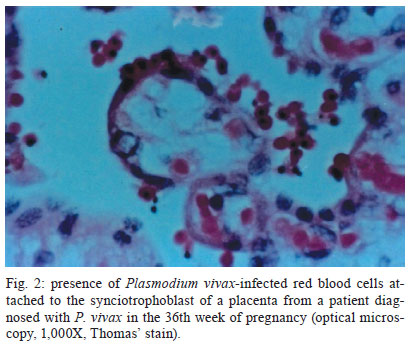
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 79-84 Original Article On cytoadhesion of Plasmodium vivax: raison d'être? Fabio TM CostaI, +; Stefanie CP LopesI; Mireia FerrerII; Juliana A LeiteI; Lorena Martin-JaularII; Maria BernabeuII; Paulo A NogueiraIII; Maria Paula G MourãoIV; Carmen Fernandez-BecerraII; Marcus VG LacerdaIV; Hernando del PortilloII, V, + IDepartamento
de Genética, Evolução e Bioagentes, Instituto de Biologia,
Universidade Estadual de Campinas, Campinas, SP, Brasil It is generally
accepted that Plasmodium vivax, the most widely distributed human malaria parasite,
causes mild disease and that this species does not sequester in the deep capillaries
of internal organs. Recent evidence, however, has demonstrated that there is
severe disease, sometimes resulting in death, exclusively associated with P.
vivax and that P. vivax-infected reticulocytes are able to cytoadhere in vitro
to different endothelial cells and placental cryosections. Here, we review the
scarce and preliminary data on cytoadherence in P. vivax, reinforcing the importance
of this phenomenon in this species and highlighting the avenues that it opens
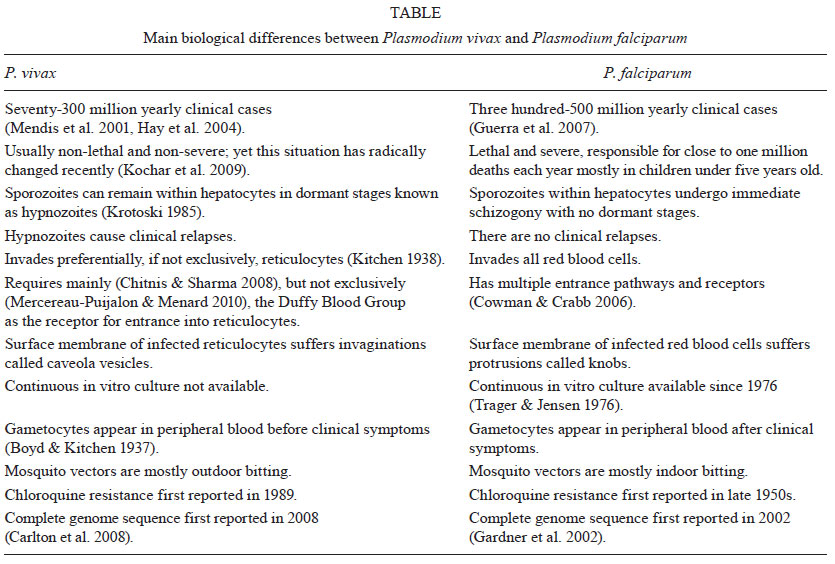
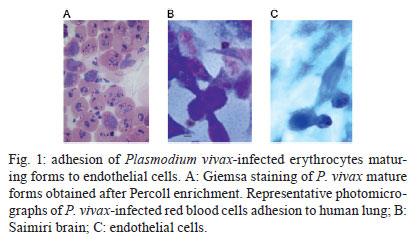
for our understanding of the pathology of this neglected human malaria parasite. Notably, the life cycle and biology of P. vivax differ from those of Plasmodium falciparum (Table). The following differences are worth highlighting: (i) P. vivax sporozoites can enter hepatocytes and remain dormant as hypnozoites, which are responsible for clinical relapses (Krotoski 1985), (ii) P. vivax preferentially, if not exclusively, invades reticulocytes (Kitchen 1938), and this is arguably the reason why parasitemias are low in natural vivax infection, (iii) gametocytes appear in the blood before clinical symptoms are manifest, which complicates and compromises clinical treatment and control of the disease (Boyd & Kitchen 1937), and (iv) mosquito vectors from areas where P. vivax is more prevalent are outdoor-biting and anthropophilic, which makes control measures, such as impregnated bed nets, of limited value. Experts therefore agree that due to this unique biology, the present tools used to control P. falciparum will not suffice for P. vivax, which reinforces the importance of understanding the biology, epidemiology and pathology of P. vivax (Mueller et al. 2009). Pathology of P. vivax - Pathogenesis in falciparum malaria is multi-factorial and involves numerous host and parasite factors. However, central to this pathogenesis are the phenomena of antigenic variation, rosetting and cytoadherence (Miller et al. 2002). Antigenic variation refers to the capacity of parasites to clonally express variant proteins at the surface of infected red blood cells (IRBCs) and this function to evade the host immune response and allow for the establishment of chronic infection (Brown & Brown 1965). Rosetting is the formation of rosettes resulting from the adhesion of erythrocytes infected with mature forms of the parasite to uninfected erythrocytes and this process has been shown to increase the microvascular obstruction of blood flow (David et al. 1988). Cytoadherence refers to the capacity of IRBC to adhere to host cells. This process has been described for the endothelium and for the placenta and it causes the disappearance of mature trophozoites and schizonts from the peripheral blood (Bignami & Bastianelli 1889, Miller et al. 1994). Remarkably, the original search for genes responsible for these phenomena identified a single subtelomeric multigene variant family named the var gene family (Baruch et al. 1995, Smith et al. 1995, Su et al. 1995). P. falciparum var genes encode proteins collectively known as PfEMP1 that contain ligand domains for different endothelial receptors, such as CD36, intercellular adhesion molecule-1 (ICAM-1) and chondroitin sulphate A (CSA). Furthermore, var gene expression is associated with various clinical syndromes of severe falciparum disease (Pasternak & Dzikowski 2009). In addition to var genes, the full genome sequence of P. falciparum has revealed additional subtelomeric multigene families that are likely involved in the pathology of malaria (Gardner et al. 2002). These data therefore indicate that expression of P. falciparum multigene families, which are clustered in subtelomeric regions, is associated with the pathology of falciparum malaria and the phenomena of antigenic variation, rosetting and cytoadherence. Studies from P. vivax patients in Sri Lanka have shown that P. vivax undergoes antigenic variation and that the mature forms have the capacity for rosetting (Udagama et al. 1987, Udomsanpetch et al. 1995). Thus, using immune sera from primary-infected or multiply infected P. vivax patients, immunofluorescence assays from different isolates have revealed that only patients with multiple infections were able to recognise several different isolates (Udagama et al. 1987). Moreover, rosetting experiments using blood and wild isolates revealed that close to 70% of the isolates showed rosetting (Udomsanpetch et al. 1995). However, approaches similar to those used for studies of cytoadhesion in P. falciparum (Hasler et al. 1993) failed to observe the cytoadhesion of P. vivax-infected reticulocytes to human umbilical vein endothelial cells, C32 melanoma cells, platelets or to purified CD36. Notably, sequence analysis of a P. vivax isolate revealed the presence of a subtelomeric multigene variant superfamily termed vir (P. vivax variant genes), which is likely involved in antigenic variation in this species (del Portillo et al. 2001). Further analyses have indicated that vir genes could have additional functions and it was hypothesised that they mediate adhesion to facilitate the establishment of chronic infection (del Portillo et al. 2004). The complete genome sequence of the Sal I strain revealed the presence of additional subtelomeric gene families (Carlton et al. 2008), which further indicated that P. vivax, like P. falciparum, has clustered multigene families at its chromosome ends. However, the relationship between these gene families and pathology has been mostly overlooked, as vivax malaria had been predominantly accepted to be clinically benign and not to cytoadhere. In the last 10 years, the severe complications observed in P. falciparum infection, which include renal failure, jaundice, acute respiratory distress syndrome, cerebral malaria, seizures, anemia, hyperparasitemia, thrombocytopenia, pulmonary edema, splenic rupture and death, have also been reported worldwide for P. vivax (Kochar et al. 2005, 2009, Baird 2007, Price et al. 2007, Alexandre et al. 2010). However, diagnosis was made in most cases using Giemsa staining, which cast reasonable doubts as to the potential for undetectable infection with P. falciparum. The use of nested polymerase chain reactions (PCRs) to exclude the possibility of mixed infection, however, has confirmed sole infection with P. vivax in some of these reports. In addition, the clinical exclusion of other syndromes, such as those caused by respiratory pathogens, human immunodeficiency virus, histoplasma, coccidians and paracoccidians, as well as the adoption of World Health Organization criteria used for severe falciparum disease, are now being adopted for the reports of new cases of severe vivax disease. In Brazil, for instance, a recent study from the city of Manaus, Amazonas, showed an increased number of hospital admissions for P. vivax-infected patients (Santos-Ciminera et al. 2007) with increased clinical severity, which were mostly related to jaundice and severe anaemia after P. falciparum or other possible causes of disease were excluded (Alexandre et al. 2010). Similar to what has been observed in Brazil, an increasing number of cases with complications have been reported worldwide (Anstey et al. 2009). These data illustrate and confirm that there is indeed an augmentation of the number of severe cases exclusively associated with P. vivax infection across different endemic regions of the world. Nevertheless, whether this severity is directly associated with changes in parasite biology, increasing resistance to antimalarial drugs or with the presence of comorbidities is still a matter of debate. Although it has long been axiomatic that P. vivax is less virulent than P. falciparum due to its lack of cytoadherence, the existence of subtelomeric variant families in the genome of P. vivax and the recent confirmed reports of severe vivax disease have led us to question the validity of this view. Moreover, in spite of studies that have documented the presence of all stages of P. vivax in the peripheral blood of infected patients, previous studies revealed that the presence of mature forms were disproportionably lower than young stages, indicating a segmentation of these mature forms somewhere else than in peripheral blood (Field et al. 1963). Cytoadherence of P. vivax-infected reticulocytes - In an attempt to elucidate whether cytoadhesion is a P. falciparum-restricted phenomenon, a recent collaborative study by our groups (Carvalho et al. 2010) demonstrated that P. vivax-IRBC (Pv-IRBCs) obtained from Brazilian patients with non-severe disease were able to cytoadhere under static or flow conditions to brain, lung (Fig. 1), placental and endothelial receptors, which have previously been implicated in the binding of P. falciparum (Beeson & Brown 2002). Under static conditions, the number per mm2 of adherent Pv-IRBC was 10-15-fold lower than that of P. falciparum-IRBC (Pf-IRBC). However, the percentage of parasites able to bind endothelial receptors under flow conditions did not differ significantly. These results differ from those previously reported in Sri Lanka (Udomsanpetch et al. 1995). One explanation for this discrepancy is that our cytoadhesion assays used percoll-enriched mature forms of the parasite at pH 6.8, whereas this other studies has used forms obtained directly from the blood at pH 7.4. To identify potential endothelial receptors involved in P. vivax adherence, CHO cells expressing different endothelial receptors were used for adhesion assays. We found greater adhesion to ICAM-1-transfected cells in comparison to CD36-transfected or untransfected cells. However, we cannot rule out the role of additional host receptors in Pv-IRBC. Moreover, adhesion of parasites to endothelial cells was significantly inhibited by the addition of soluble CSA, although treatment of the same cells with chondroitinase-ABC (CaseABC) did not affect adhesion (Carvalho et al. 2010). These results indicated that the CSA backbone was not directly involved in parasite adhesion. As CSA is highly negatively charged due to the presence of sulphate groups, the inhibition of cytoadhesion may be sustained by a charge interaction. Indeed, it has been shown that the binding of several P. falciparum VAR2CSA domains to CSA at various salt concentrations is not different from their binding to non-placental domains, which demonstrates that binding to CSA is highly dependent on ionic strength (Resende et al. 2009). We hypothesised that Vir proteins, due to their variant nature, sub-cellular localisation and genome organisation, could mediate P. vivax cytoadherance. By using two different antibodies raised against two different Vir proteins, the specific inhibition of the IRBC-endothelial cell interaction was demonstrated (Carvalho et al. 2010). To perform functional binding assays in the absence of a continuous in vitro culture system for P. vivax, 3D7 P. falciparum transgenic lines expressing different Vir proteins, motifs, protein domains and secondary structures have been generated (M Bernabeu & C Fernandez-Becerra, unpublished observations). Notably, one of these transgenic lines that expressed Vir proteins on the surface of the IRBC mediated cytoadherence to ICAM-1, VCAM and E-selectin. Moreover, two polyclonal, mono-specific anti-Vir antibodies significantly inhibited this adhesion. Together, these data further support our findings and indicate that P. vivax-infected reticulocytes can cytoadhere to a variety of endothelial receptors and that this adherence is at least partially mediated specifically by Vir proteins. Cytoadherence and severe vivax disease - The recent findings that infection with P. vivax can lead to severe complications in endemic areas worldwide (Kochar et al. 2005, Lomar et al. 2005, Tjitra et al. 2008) have led the scientific community to search for the mechanisms of pathogenesis related to this infection, which have been neglected thus far. One of the major arguments against the severity of P. vivax infection was based on the lack of cytoadherence and the formation of knobs by this parasite. However, robust evidence points to the existence of severe falciparum disease without the presence of the aforementioned mechanisms, which therefore suggests that these are not the sine qua non for severity (Maegraith 1948). As reviewed above, cytoadhesion was demonstrated for P. vivax using an in vitro approach (Carvalho et al. 2010) and unpublished data on adhesion to the placenta (FTM Costa & MVG Lacerda, unpublished observations) (Fig. 2), lungs and spleen (MVG Lacerda & H del Portillo, unpublished observations) have recently been obtained. When IRBCs are found inside of blood vessels in human tissues, there is no way to confirm that cytoadhesion has taken place, and this possibility should also be taken into account for previous studies of P. falciparum-infected tissues. There is not much information available on autopsies related to vivax infection, which could lend support to cytoadherence during severe vivax disease. As reviewed elsewhere (Anstey et al. 2009), however, early XX century autopsies for which P. vivax diagnosis was based on thick blood smears reported the presence of intracapillary masses of swollen, infected erythrocytes and pigment (Billings & Post 1915). In addition, these autopsies reported the frequent presence of mature malaria parasites within a red blood cell taking up the entire lumen and being in immediate contact with endothelial cells in some vessels in the brain, as well as an unusually large number of infected red cells and young plasmodia and a large amount of pigment in intestinal submucosal vessels (Bruetsch 1932). In these cases, however, comorbidities and mixed infection with P. falciparum were not properly discarded. More recently, one autopsy from India, which had confirmed P. vivax monoinfection by PCR, found congestion of alveolar capillaries by monocytic infiltrates and diffuse damage to alveolar membranes consistent with acute respiratory distress syndrome. Parasites were observed in lung tissue as well as in other organs without lesions (Valecha et al. 2009). Despite the fact that this is evidence from a single case, this resembles the findings of cytoadhesion in P. falciparum pulmonary disease (Corbett et al. 1989). From measurements of gas transfer and lung volume in non-severe vivax patients, the progressive alveolar-capillary dysfunction following treatment was found to be consistent with the greater inflammatory response induced by this parasite (Anstey et al. 2007). As a major complication of severe vivax disease, the pathogenesis of respiratory distress requires further investigation into the cytoadhesion that takes place, as suggested by studies of in vitro adhesion and endothelial activation of lung microvasculature. In fact, the host inflammatory response, the endothelial Weibel-Palade body release, and the plasma concentrations of angiopoietin-2, ICAM-1, E-selectin, interleukin (IL)-6 and IL-10 are greater in patients with vivax compared to P. falciparum malaria (Yeo et al. 2010). As the addition of lipopolysaccharide was found to increase the adhesion of IRBCs to human lung endothelial cells in vitro (Carvalho et al. 2010), it is possible that secondary bacterial infections may release endotoxins that could contribute to the enhanced endothelial activation and cytoadhesion. Thus, coinfections and pathogenesis during vivax infection require further investigation. The role of platelets in cytoadhesion is well-documented for severe falciparum malaria. Here, platelets participate in strengthening the adhesion of IRBCs to CD36 (known as glycoprotein IV in platelets), which most likely explains their accumulation in brain microvessels (Grau et al. 2003, Wassmer et al. 2004). Platelets can also participate in tumor necrosis factor (TNF)-induced microvascular pathology (Lou et al. 1997, Wassmer et al. 2006) and platelet-derived microparticles seem to play an important role in endothelial activation (Combes et al. 2006). Despite strong evidence that vivax disease triggers greater thrombocytopenia than does falciparum disease (Kochar et al. 2010), there are no data available on the ability of these particles to enhance vivax cytoadhesion. As reviewed elsewhere (Clark & Alleva 2009), sequestration in the bone marrow has been shown to cause the dyserythropoiesis observed in P. falciparum malaria (Wickramasinghe et al. 1987). Dyserythropoiesis was demonstrated in the bone marrow during P. vivax infection, where cytoadhesion was not thought to play a role (Wickramasinghe et al. 1989). As a result, the idea of cytoadhesion as the cause of severe anaemia was abandoned and the presence of increased TNF levels was shown to induce dyserythropoiesis in both species (Clark & Chaudhri 1988). The presence of P. vivax parasites in the bone marrow prompts the need for clarification on its implications for disease pathogenesis (Lacerda et al. 2008). Perspectives and concluding remarks - Demonstration of the cytoadherence of P. vivax opens new investigational lines of research. First, proteomic analysis of adhesion proteins and Vir proteins on the surface of infected reticulocytes should further the identification of new parasite ligands. In addition, microscopic analyses from autopsies where coinfections have been ruled out will reinforce the presence of parasite cytoadherence. The role of cytoadherence to the spleen, lungs and placenta in the establishment of chronic infection will further reveal the molecular basis of pathology in P. vivax malaria and the demonstration of cytoadhesion in other tissues, such as the kidneys and bone marrow will confirm existing observations of severe disease and the association between cytoadhesion and severe disease syndromes. Furthermore, technological breakthroughs in noninvasive imaging will demonstrate whether P. vivax-infected reticulocytes adhere to different organs in vivo and whether this adhesion is associated with severe vivax malaria. Ultimately, understanding the factors that determine P. vivax-associated morbidity and severe disease will contribute to the control, elimination and eradication of malaria worldwide (Bassat & Alonso 2011). References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11143t1.jpg] [oc11143f2.jpg] [oc11143f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}