 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 85-90 Original Article Surface-expressed enolases of Plasmodium and other pathogens Anil Kumar Ghosh; Marcelo Jacobs-Lorena+ Department of Molecular
Microbiology and Immunology and Malaria Research Institute, Johns Hopkins Bloomberg
School of Public Health, 615 N Wolfe Street, Baltimore, 21205 MD, USA
Received 22 April
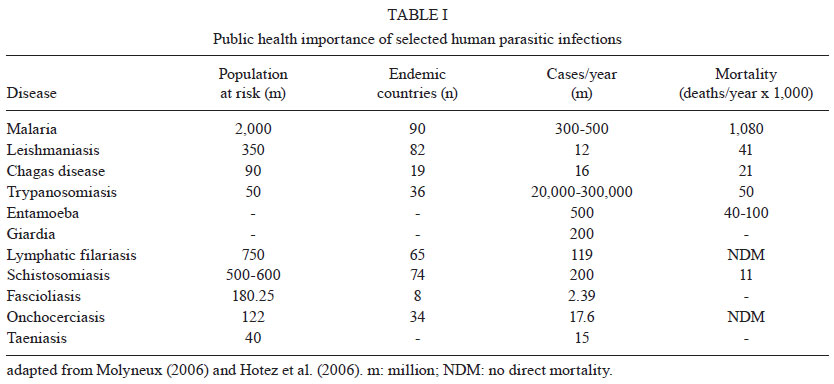
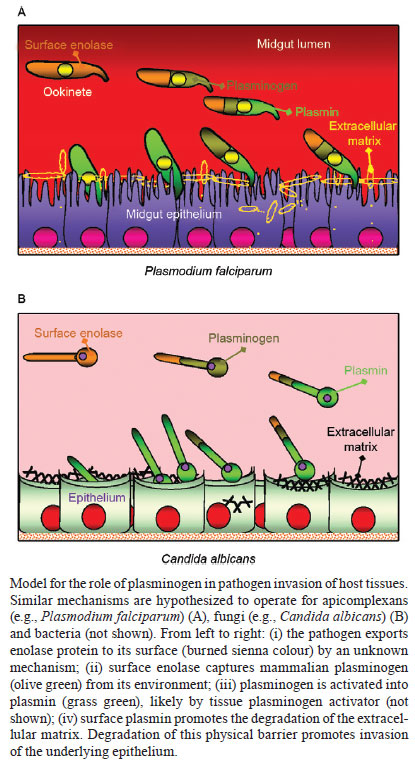
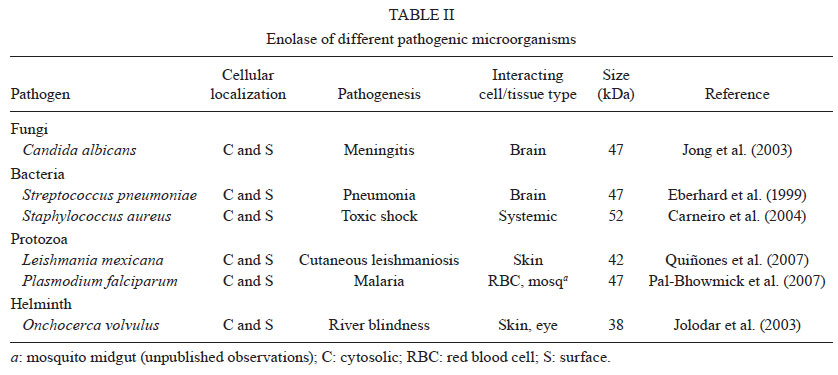
2011 Code Number: oc11144 Abstract Enolase is the eighth enzyme in the glycolytic pathway, a reaction that generates ATP from phosphoenol pyruvate in cytosolic compartments. Enolase is essential, especially for organisms devoid of the Krebs cycle that depend solely on glycolysis for energy. Interestingly, enolase appears to serve a separate function in some organisms, in that it is also exported to the cell surface via a poorly understood mechanism. In these organisms, surface enolase assists in the invasion of their host cells by binding plasminogen, an abundant plasma protease precursor. Binding is mediated by the interaction between a lysine motif of enolase with Kringle domains of plasminogen. The bound plasminogen is then cleaved by specific proteases to generate active plasmin. Plasmin is a potent serine protease that is thought to function in the degradation of the extracellular matrix surrounding the targeted host cell, thereby facilitating pathogen invasion. Recent work revealed that the malaria parasite Plasmodium also expresses surface enolase, and that this feature may be essential for completion of its life cycle. The therapeutic potential of targeting surface enolases of pathogens is discussed. Key words: Plasmodium - cell surface enolase - pathogenicity The global burden of parasitic diseases is a problem of enormous proportions, especially in developing countries (Table I). The highest burden is inflicted by malaria, with the number of people at risk approaching two billion and expected to keep increasing (Snow et al. 2005). Malaria is most severe in the sub-Saharan Africa where a child dies every 40 s and imposes an annual economic burden of $12 billion (Rugemalila et al. 2007). To survive, obligate intracellular pathogens need to invade their host tissues. Within their target cells the microbes are safe to propagate and prepare for transmission to the next host. Pathogens from bacteria to helminths have developed a variety of mechanisms to invade host tissues. Several bacteria develop fimbria or different types of pilli, for the initial contact with their host cells. Other protozoa develop pseudopodia, which are undulating membranous flagella, for increased penetration efficiency. Helminthic parasites use a variety of cutting appendages and suckers for anchoring and invasion. In addition to the above-mentioned structural modifications, several biochemical arsenals have developed during the course of evolution, including adhesion molecules and cellular receptors (Pizarro-Cerdá & Cossart 2006). A number of pathogenic organisms display specialized proteins on their cell surface to assist in invasion, one of the best characterized being the glycolytic enzyme enolase. Cell surface enolase efficiently captures plasminogen from the surrounding environment which is subsequently converted into plasmin, a strong serine protease that facilitates the invasion process (Lottenberg et al. 1994, Ehinger et al. 2004). Here we review what is known about the role played by surface enolase in Plasmodium and other pathogens. Enolase and plasminogen - Enolase is an ancient and ubiquitous metalloenzyme. It catalyzes the reversible dehydration of 2-phosphoD-glycerate into P-enolpyruvate during glycolysis. Eubacteria and archaebacteria have a single enolase gene and therefore multiple molecular forms (isoforms) are not observed. Overall, enolase sequences are highly conserved, showing roughly 40% amino acid identity across eukaryotes, eubacteria and archaebacteria although a large number of insertion and deletions occur across phyla. Some of the insertions and deletions have been shown to be highly plastic (Keeling et al. 2004). In yeast and most higher organisms, enolase subunits assemble into dimers while in some eubacteria (e.g. Bacillus subtilis and Thermotoga maritima) the protein is octameric. Plasmodium has a single enolase gene in chromosome 10.92% A + T content (Read et al. 1994). The protein has 446 amino acids residues (48.7 kDa), similar in length to enolases from other eukaryotes. Plasmodium enolase has a common ancestry with apicomplaxan and plants and may have derived from their plastid endosymbionts (Read et al. 1994, Hannaert et al. 2000). Plasmodium enolase carries a EWGWS pentapeptide insertion (residues 104-108) that is immunogenic and may be exposed on the surface of the parasites (Vora et al. 2009). It is unclear how enolase is exported to the surface of the parasite. The absence of a signal peptide indicates that enolase does not transit through the endoplasmic reticulum but it has been suggested that enolase enters the classical secretory pathway by a previously unidentified mechanism (López-Villar et al. 2006). Plasminogen is a 92 kDa proenzyme that requires cleavage at its C-terminal region (Ponting et al. 1992) for activity. Activation of plasminogen into active plasmin involves cleavage of the Arg561-Val562 bond yielding an N-terminal A (heavy) chain which remains linked by two disulphide bonds to the smaller C-terminal B (light) chain. The B chain contains the protease-active residues (Ponting et al. 1992). The plasminogen N-terminal region contains five 80-amino-acids-long Kringle domains. Plasminogen binds strongly and specifically to surface enolase via interaction of its Kringle domains with the C-terminal or internal lysine motifs of enolase (Derbise et al. 2004). Plasmodium surface enolase and midgut invasion - For transmission to occur, Plasmodium needs to complete a complex life cycle within its mosquito vector. The cycle begins with the rapid differentiation of gametocytes acquired with the infected blood meal into male and female gametes, followed by fertilization and differentiation of the resulting zygotes into motile ookinetes. The ookinete must then cross two physical barriers: the peritrophic matrix (Shao et al. 2001) and the midgut epithelium (Ghosh et al. 2000). We recently found that enolase is exported to the surface of ookinetes but not of gametocytes, gametes and zygotes (AK Ghosh et al., unpublished observations). Ookinete surface enolase appears to perform two separate functions: (i) bind to a mosquito midgut epithelial receptor for invasion of the midgut and (ii) recruit plasminogen (present in the ingested blood) via enolase's internal lysine motif (DKSLVK). Recruitment of plasminogen to the ookinete surface is essential, as depletion of plasminogen from the blood meal completely aborts parasite development in the mosquito. Replenishment of the plasminogen-depleted blood with recombinant wild type plasminogen restores mosquito infection, while addition of a plasminogen point mutant in plasmin's active site does not restore infection (AK Ghosh et al., unpublished observations). These results suggest that not only is plasminogen required, but that its conversion to active plasmin is essential. We speculate that plasmin functions in degradation of the extracellular matrix (glycocalyx) that covers the mosquito midgut epithelium (Zieler et al. 2000) (A in Figure). Sato et al. (2000) found that patients infected with Plasmodium falciparum and Plasmodium vivax carry high anti-enolase antibody titres and concluded that enolase could be used for the immunodiagnosis of malaria. Roth et al. (1988) found that the total amount of enolase in parasitized red blood cells (RBCs) increases about 15-fold during infection. Recently, we found that enolase also inserts on the surface of the RBCs infected with the human parasite P. falciparum (unpublished observation). The role played by this RBC surface enolase is not known. Enolase-mediated cell invasion is a common theme among pathogenic microorganism - A brief summary of the occurrence of cell surface enolase across different phyla and their potential role in pathogenesis is provided in Table II. Candida albicans can cause superficial infections of the oral and vaginal mucosa and in some cases causes life-threatening disseminated disease (Cutler 1991). C. albicans may also invade the central nervous system resulting in devastating meningitis (Zhang et al. 1999). To reach the central nervous system C. albicans must cross the blood-brain barrier by traversing the endothelial lining of the brain capillaries. It was found that Candida binds both plasminogen and plasmin in a specific and saturable manner. Plasminogen binding to the Candida surface, which is mediated by surface enolase, greatly enhances its affinity for streptokinase, an enzyme involved in plasminogen activation to plasmin. The net result is enhanced fibrinolysis and increased ability to cross an in vitro blood-brain barrier system (Jong et al. 2003) (B in Figure). Streptococcus pneumoniae is the causative agent of bacterial meningitis. S. pneumoniae can degrade extracellular matrix components thus breaching the blood-brain barrier and penetrating brain tissue to cause severe disease. The S. pneumoniae enolase gene encodes a 47 kDa protein with no signal sequence or transmembrane domain and as for other pathogens it is not known how this protein reaches the cell surface and remains attached to it. The S. pneumoniae enolase protein is octameric and has two plasminogen-binding motifs, one at the C-terminal end and the other internal, from amino acid 248-256 (Ehinger et al. 2004). In an elegant experiment where reconstituted basement membrane (Matrigel) was used to test the penetration process, it was observed that bacteria only invade the basement membrane when in medium containing plasminogen and when surface bound plasminogen was cleaved by tissue plasminogen activator (tPA). In the absence of plasminogen bacteria could not penetrate even after 21 h incubation. The results suggest a potential role of surface-associated plasminogen in bacterial penetration of basement membranes and extracellular matrices (Eberhard et al. 1999). Staphylococcus aureus is a highly invasive bacterium that causes a large number of the hospital infections throughout the world, having a major impact on human morbidity and mortality. S. aureus can colonize the mucosa of airways and also has a propensity for targeting bone tissues, thereby causing bone disease, especially osteomyelitis (Wheat 1985). The bacterium produces a 52 kDa protein with high homology with enolase that is transported to the cell wall surface by an unknown mechanism. This 52 kDa protein binds to laminin and might function by first allowing S. aureus to adhere to the extracellular matrix, followed by plasminogen activation and laminin degradation around the penetration area (Carneiro et al. 2004). Streptococcus pyogenes is responsible for a wide variety of human disease that range from infections of the throat (pharyngitis), skin (impetigo) and degeneration of underlying tissue (necrotizing fasciitis or flesh-eating disease), to a total toxic shock syndrome, rheumatic fever and acute glomerulonephritis (Pancholi & Fischetti 1998). S. pyogens expresses a protein of about 45 kDa called streptococcal surface enolase on its surface. It bears a C-terminal lysine residue which binds to the Kringle domain of plasminogen thus promoting the invasion process (Derbise et al. 2004). Leishmania spp are protozoan parasites that are the cause of cutaneous and visceral leishmaniasis. Leishmania mexicana expresses a 47 kDa enolase that localizes mainly to the cytoplasm but a significant proportion is found facing the external surface of the cell (Quiñones et al. 2007). Leishmania surface enolase binds plasminogen (Avilan et al. 2000, Calcagno et al. 2002) and this interaction contributes to the virulence of the parasite (Maldonado et al. 2006). An anti-enolase antibody interferes with plasminogen binding, making this a vaccine candidate antigen. Human onchocerciasis is caused by the filarial parasite Onchocerca volvulus and is a major cause of skin pathology and eye lesion, including blindness (river blindness). An estimated 18 million people are infected and about 120 million people are at risk of infection mainly in tropical Africa. The parasite is transmitted by female black flies of the genus Simulium. The incubation period in man is about one year. The infected larvae remain localized in the skin and grow into adults producing large numbers of microfilariae that are the form transmitted by black flies and the main cause of disease symptoms. O. volvulus encodes an enolase of about 38 kDa. The protein was observed to be localized to the afibrillar portion of the body wall muscle and to the uterus. It is not known how the protein is transported to the outer cell surface (Jolodar et al. 2003). Therapeutic potential of targeting cell surface enolase - Previous mouse infection and in vitro experiments indicated that binding of plasminogen to pneumococci, and in particular to surface-exposed enolase, is important for in vivo virulence and plasmin-mediated degradation of the extracellular matrix (Bergmann et al. 2003, 2005). The presence of anti-pneumococcal antibodies in sera from young children with and without a history of pneumococcal contact has been compared (Adrian et al. 2004). No significant correlation between antibody titres and infection was observed. Since enolase is a conserved glycolytic enzyme, it was concluded that anti-enolase antibodies in young children probably represented cross-reactive epitopes to commensal bacteria (Adrian et al. 2004). Epitope accessibility on viable bacteria is of critical importance in the development of protective antibodies to bacterial infections. The epitope of the anti-enolase mAb (245, C-6) was mapped to residues 55DKSRYGGLG63 in the N-terminal domain of S. pneumoniae enolase (Ehinger et al. 2004). Crystal structure of the protein indicated that these residues are surface-exposed on the enolase octamer. Immuno-electron microscopy and flow cytometric studies revealed that the abundance of surface-exposed enolase of in vitro-grown pneumococci is relatively low. However, pneumococcal colonization is a multifactorial process which also might involve changes in expression of the enolase protein under in vivo conditions (Kolberg et al. 2006). Despite the low amount of enolase protein on in vitro-grown pneumococci, the efficiency of plasminogen binding is high and this amount is sufficient to contribute to pneumococcal pathogenesis as shown previously by Bergmann et al. (2003) and Kolberg et al. (2006). Streptococcus suis serotype 2 (SS2) is a porcine and human pathogen that can cause meningitis, endocarditis and septic shock in people who handle swine and its products (Trottier et al. 1991). Mice immunized with anti-recombinant enolase gave rise to a high titre humoral response resulting in an 83% protection after challenge with the SS2 bacterium (Zhang et al. 2009). Dental caries is a prevalent chronic human infection caused by Streptococcus sorbinas and Streptococcus mutants. When a formulation of recombinant sorbinas enolase in alum was administered to the oral cavity of rats, it elicited a high humoral antibody response while reducing enamel loss and dental caries (Dinis et al. 2009). Immunofluorescence as well as electron microscopic examinations revealed that P. falciparum enolase localizes on the merozoite surface. Mice immunized with recombinant P. falciparum enolase showed protection against a challenge with a lethal strain of Plasmodium yoelii suggesting that enolase is a potential protective antigen (Pal-Bhowmick et al. 2007). In a separate set of experiments mice were immunized with recombinant P. falciparum enolase and then infected with Plasmodium berghei. Mosquitoes feeding on the infected immunized mice had a significant lower number of oocysts, suggesting that enolase could be used as an antigen for a transmission blocking vaccine (unpublished observations). Perspectives - The life cycle of pathogens frequently involves crossing of physical barriers such as extracellular matrices and tissue epithelia. This is true for pathogen development in their mammalian as well as invertebrate hosts. Mammals make use of the potent protease plasmin, to regulate its clotting system. Plasminogen, the plasmin precursor, binds to fibrin via its Kringle motifs and this binding increases its susceptibility to activating enzymes such as tPA. It is interesting that such a wide variety of pathogens, from bacteria to multicellular organisms, opted to recruit the plasminogen-plasmin system of their mammalian hosts to their own advantage. An additional challenge for the pathogen is that plasminogen needs to be captured on the pathogen's surface which is devoid of fibrin. To this end, pathogens evolved a mechanism to export one of their metabolic enzymes enolase or a related protein, to the cell surface, as these proteins carry a lysine motif that physically interacts with the Kringle motifs of plasminogen. The genome of these pathogens frequently has a single enolase gene whose protein product has to accomplish two important functions: act (i) as a metabolic enzyme and (ii) as a surface anchor. What mechanisms operate to provide such different functions to the same protein is not known for any organism. After synthesis on polyribosomes, what determines whether the protein (enolase or enolase-like) remains in the cytoplasm or is exported to the surface? As the protein does not have a recognizable signal sequence, by what mechanism(s) does it reach the surface? As the protein does not have a recognizable trans-membrane domain, by what mechanism(s) does it stay attached to the pathogen surface? In any event, it is interesting that such a broad range of organisms (convergently?) evolved the same strategy to capture plasminogen on their surface. Additional outstanding questions include the mechanisms of plasminogen activation. As plasminogen binding to fibrin makes it more susceptible to its activating enzymes, it is reasonable to speculate that the same is true for plasminogen binding to enolase on the pathogen surface. However, not much is known about which enzymes actually catalyze the conversion of plasminogen to plasmin on the pathogen surface. In case of Plasmodium, our laboratory has preliminary evidence that tPA binds to the surface of the mosquito midgut epithelium, suggesting the hypothesis that activation of plasminogen on the ookinete surface actually occurs at the time of ookinete-midgut epithelium contact. Another outstanding question is the nature of the protein substrates of surface plasmin during tissue invasion. In mammals, the principal activity of plasmin is the degradation of fibrin. However, plasmin also degrades non-collagenous extracellular matrix proteins such as laminin (Carneiro et al. 2004). In an in vitro experiment it was also found that Hemophilus influenzae can degrade laminin and fibronectin (Virkola et al. 1996). What are the substrates that are cleaved by plasmin during pathogen invasion? The answer to this question is of obvious importance as such knowledge could be explored to devise novel approaches for disease control. Acknowledgements To Johns Hopkins Malaria Research Institute and P. falciparum core facilities, for help with mosquito rearing and parasite cultures, and to Dr Clive Schiff, for his comments on the epidemiology of parasitic diseases. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11144f1.jpg] [oc11144t2.jpg] [oc11144t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}