
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 106, Num. s1, 2011, pp. 91-104
|
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 91-104
Original Article
Malaria-related
anaemia: a Latin American perspective
Juan Pablo QuinteroI,
VII; André Machado SiqueiraII , III; Alberto TobónIV;
Silvia BlairIV; Alberto MorenoV, VI; Myriam Arévalo-HerreraI,
VII; Marcus Vinícius Guimarães LacerdaII, III;
Sócrates Herrera ValenciaI, VII, +
ICaucaseco
Scientific Research Center, Cali, Colômbia
IIFundação de Medicina Tropical Dr Heitor Vieira Dourado,
Manaus, AM, Brasil
IIIUniversidade do Estado do Amazonas, Manaus, AM, Brasil
IVUniversidad de Antioquia, Medellin, Colombia
VEmory Vaccine Centre, Yerkes National Primate Research Centre, Atlanta,
GA, USA
VIDivision of Infectious Diseases, Department of Medicine, Emory
University, Atlanta, GA, USA
VIICentro Latino Americano de Investigación en Malaria, Cali,
Colombia
+ Corresponding author: sherrera@inmuno.org
Received 4 April
2011
Accepted 4 July 2011
Code Number: oc11145
Abstract
Malaria is the
most important parasitic disease worldwide, responsible for an estimated 225
million clinical cases each year. It mainly affects children, pregnant women
and non-immune adults who frequently die victims of cerebral manifestations
and anaemia. Although the contribution of the American continent to the global
malaria burden is only around 1.2 million clinical cases annually, there are
170 million inhabitants living at risk of malaria transmission in this region.
On the African continent, where Plasmodium falciparum is the most prevalent
human malaria parasite, anaemia is responsible for about half of the malaria-related
deaths. Conversely, in Latin America (LA), malaria-related anaemia appears to
be uncommon, though there is a limited knowledge about its real prevalence.
This may be partially explained by several factors, including that the overall
malaria burden in LA is significantly lower than that of Africa, that Plasmodium
vivax, the predominant Plasmodium species in the region, appears
to display a different clinical spectrus and most likely because better health
services in LA prevent the development of severe malaria cases. With the aim
of contributing to the understanding of the real importance of malaria-related
anaemia in LA, we discuss here a revision of the available literature on the
subject and the usefulness of experimental animal models, including New World
monkeys, particularly for the study of the mechanisms involved in the pathogenesis
of malaria.
Key words:
Plasmodium falciparum - Plasmodium vivax - malaria - anaemia -
haemoglobin - Latin America
Malaria is the
most important parasitic disease worldwide, causing 225 million clinical cases
and an estimated 781,000 fatalities annually, and represents a major global
public health problem (WHO 2010). Although four Plasmodium species have
been classically responsible for human malaria, Plasmodium falciparum has been the most prevalent overall, particularly in Africa (86%). On this continent,
the greatest mortality is associated with cerebral malaria and severe anaemia,
mostly in children less than five years of age, in malaria holoendemic areas
(Guerra et al. 2010). In sub-Saharan Africa, pregnant women are also at higher
risk of cerebral malaria and anaemia, which are consequently the major causes
of perinatal morbidity and mortality. Both of these malarial complications are
responsible for a great number of spontaneous abortions, stillbirths, premature
deliveries and low birth weight (Dicko et al. 2003).
Because of the
higher P. falciparum global prevalence, morbidity and mortality, most
research efforts on malaria pathogenesis have been focused on this species (Akhwale
et al. 2004). However, Plasmodium vivax represents the second most prevalent
species, responsible for an estimated 25-40% of the reported malaria clinical
cases (Westenberger et al. 2010). Until recently, there was a mistaken belief
that P. vivax was always a benign disease, however, there is growing
body of evidence of the high prevalence of severe and complicated P. vivax
malaria cases, including severe anaemia (Genton et al. 2008, Tjitra et al. 2008,
Kochar et al. 2009, Alexandre et al. 2010, Andrade et al. 2010). Additionally,
it is likely that the incidence of anaemia may be higher than what is currently
diagnosed. Multiple factors indicate that the public health relevance of P.
vivax may be more significant than was traditionally thought: (i) P.
vivax has a wider geographical range - potentially exposing more people
to the risk of infection, (ii) it is less amenable to control and (iii) most
importantly, infections with P. vivax can cause severe clinical syndromes
(Tjitra et al. 2008).
Approximately 170
million people live at risk of P. vivax and P. falciparum transmission
in 21 countries in Latin America (LA) and the Caribbean (Guerra et al. 2008,
2010). Nearly 60% of the malaria cases in the Americas are reported from Brazil
and the other 40% are reported from Colombia (14.2%), Peru (8.8%), Venezuela
(5.4%), Bolivia (1.9%) and Ecuador (1.1%). Caribbean cases include those reported
in Haiti (2.8%). Central American countries report the occurrence of malaria
cases as Guatemala (3.8%), Panama (0.4%) and Honduras (1.5%). In terms of malaria
species distribution, 74% of infections are caused by P. vivax, 25% by
P. falciparum and < 0.01% by Plasmodium malarie. All the species
together contributes a mortality estimated in less than 0.1% to the overall
mortality caused by malarial infections (WHO 2009).
The benefits of
a detailed knowledge of P. vivax transmission and its clinical burden
are identical to those of P. falciparum. The development of the Malaria
Atlas Project has shown that the global mapping of malaria is a fundamental
step to (i) understand the epidemiology of the disease on a global scale, (ii)
appraise the equity of global financing for malaria control and (iii) set the
basis for disease burden estimation. Although significant progress has been
made in P. falciparum mapping, in the case of P. vivax, such maps
have been developed only recently, making any strategic planning in LA more
difficult (Guerra et al. 2010).
To elucidate the
molecular mechanisms involved in the pathogenesis of malaria-induced anaemia,
this review address the malarial anaemia immune pathogenesis process and the
relevance of currently available experimental animal models, particularly New
World monkeys, which are susceptible to human malaria parasites (Alexandre et
al. 2010, Andrade et al. 2010).
Epidemiology
of malaria-related anaemia in LA - It has been estimated that nearly 50%
of the population, distributed in 21 countries of the American continents, is
exposed at some level to the risk of malaria transmission (Gusmao 1999). Of
these countries, Brazil and Colombia experience greater than 60% of the malaria
cases. In LA, malaria exhibits epidemiological characteristics that appear to
be particular to the region. There is an extraordinary parasite genetic differentiation
due to bio-geographic barriers such as the Andean ridge, which separates endemic
areas on the Pacific coast of the region from those in the Amazon and Orinoco
Basins.
In the case of
P. falciparum, there is substantial spatial and temporal heterogeneity
in the proportion of infections caused by each parasite population (Cortese
et al. 2002, McCollum et al. 2007). This has resulted in the isolation of parasite
populations and limited dispersion of mutations, such as those associated with
anti-malarial drug resistance in P. falciparum populations, which may
have consequences for malaria control strategies. Drug resistance associated
mutations that are common in the Amazon and Orinoco Basins have not been introduced
to the Pacific coast region (Bacon et al. 2009, Corredor et al. 2010). Additionally,
there is a dense forest, known as the Darien gap, which geographically separates
Central America from South America and maintains parasite populations that harbour
significantly different drug sensitivity profiles (Restrepo-Pineda et al. 2008).
Interestingly, communities in these distinct geographical areas correspond to
highly diverse ethnic groups (e.g., African descendants, native Indians and
mestizos) with critical genetic differences in their Duffy (Fy) blood groups,
which influence P. vivax malaria susceptibility (Hadley & Peiper
1997). As a consequence of this complexity, but also because of the limited
resources traditionally allocated to malaria research in this region, the epidemiology
of malaria, including the prevalence of anaemia in these areas, is still poorly
understood.
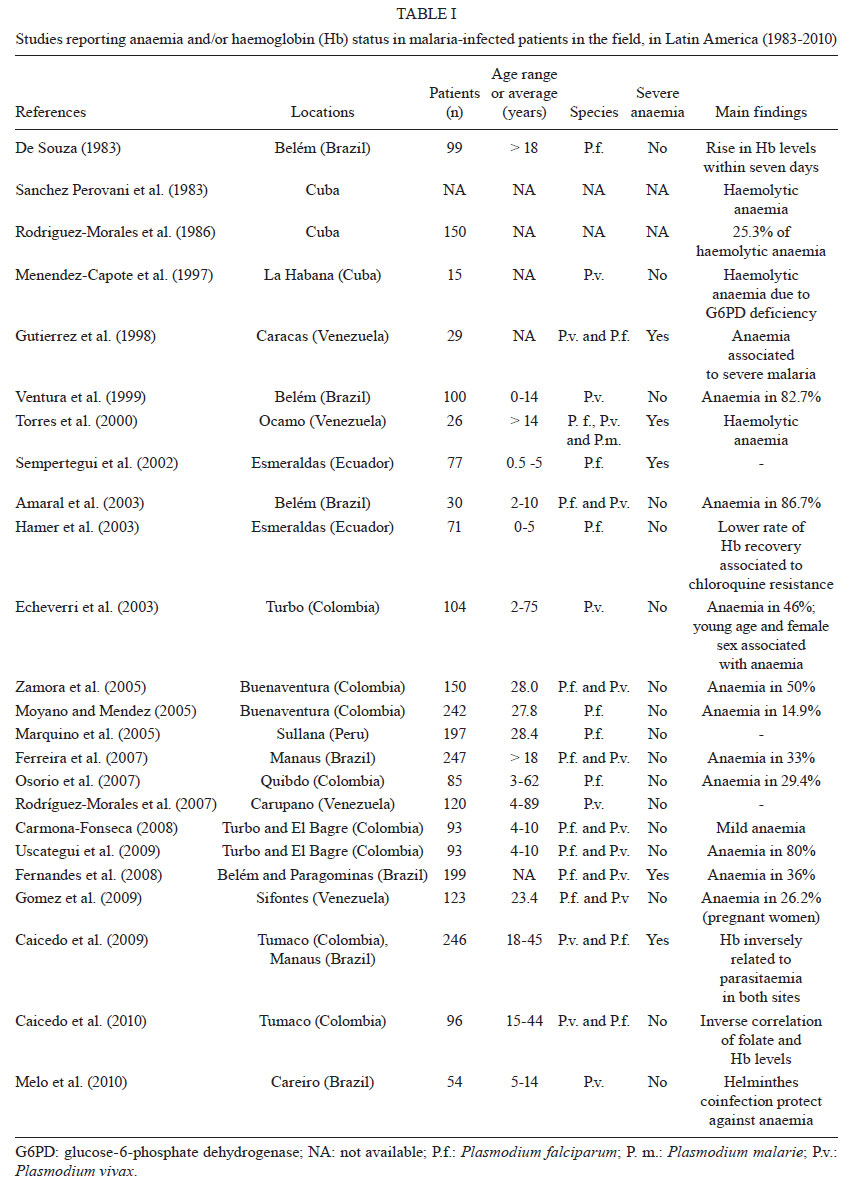
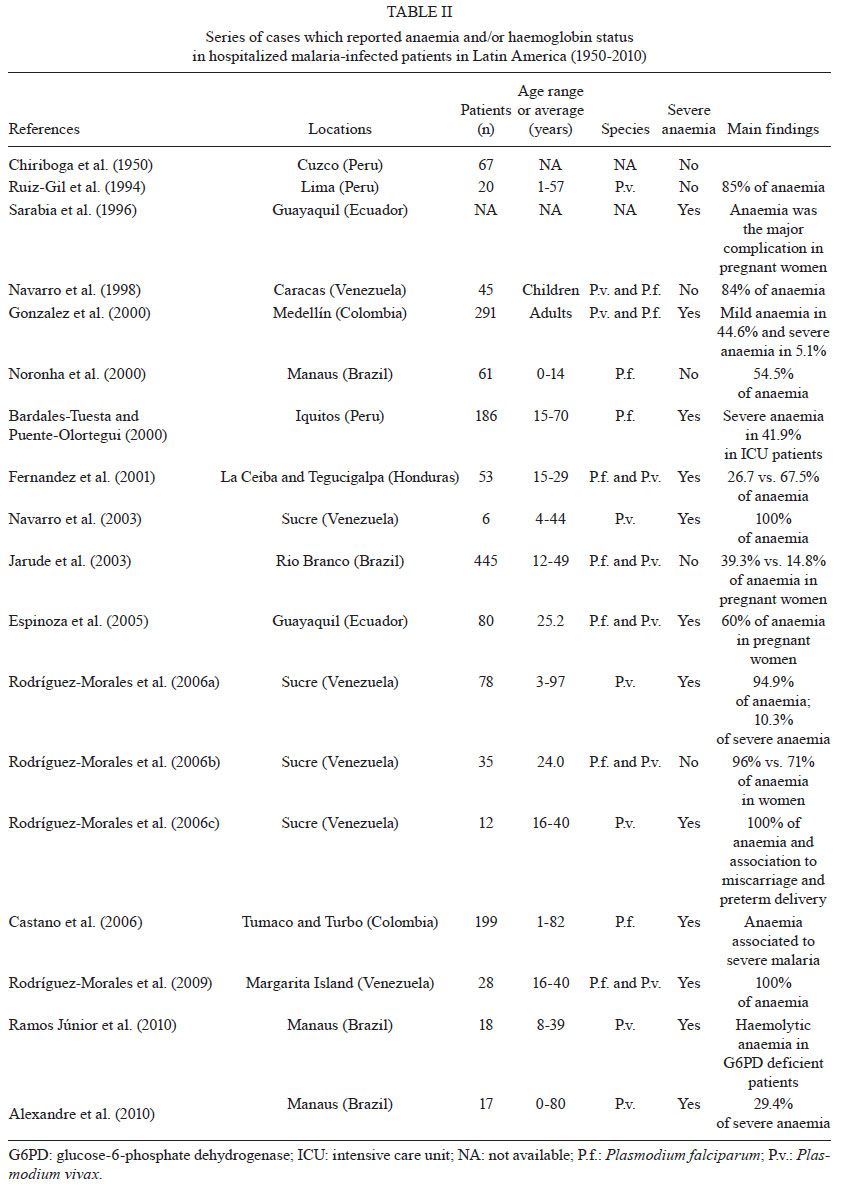
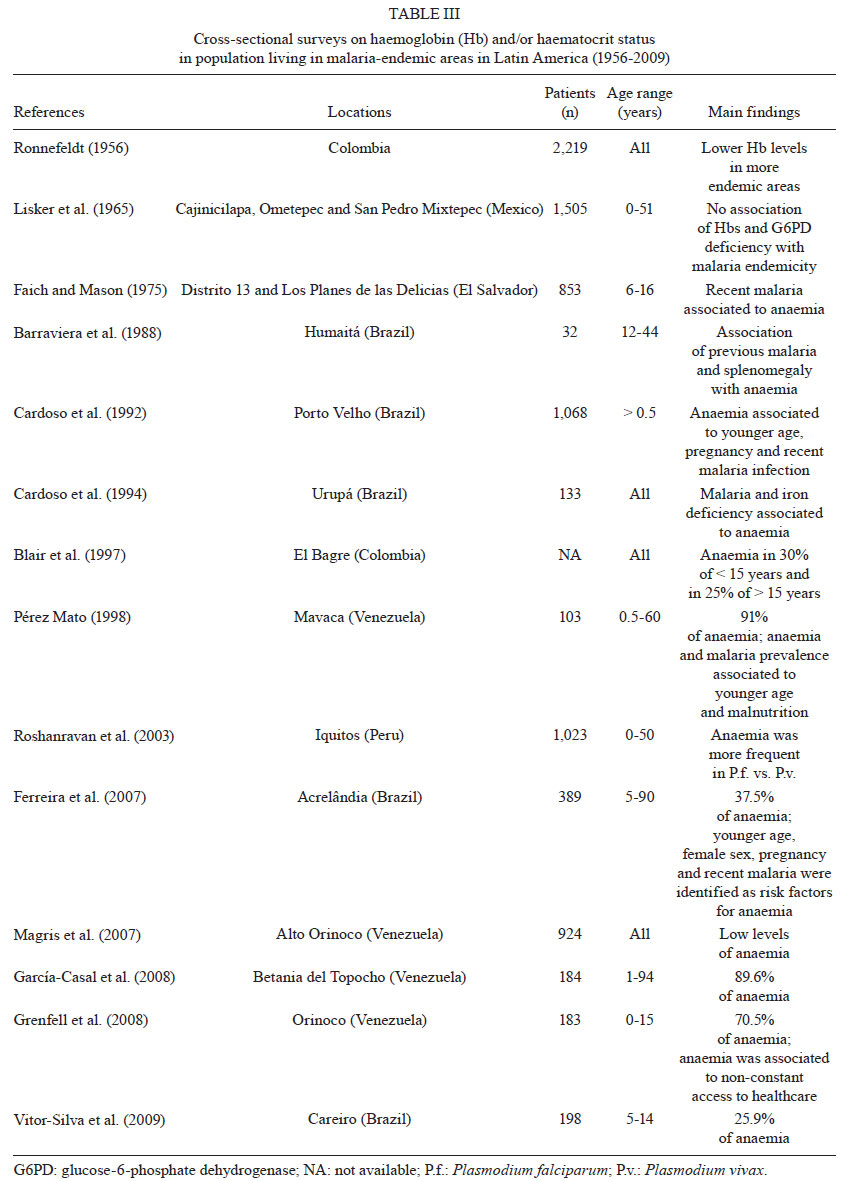
An exhaustive search
for studies addressing malaria-related anaemia in the 21 countries of LA known
to have malaria transmission has indicated that only 56 studies have been published
in this period. This represents the work spanning the more than 60 years since
the initiation of the Malaria Eradication Campaign. Twenty-four of the studies
reported anaemia and/or information on haemoglobin (Hb) levels in community-based
malaria patients (Table I), 18 studies were based
on reports of hospitalised malaria patients (Table
II) and 14 corresponded to cross-sectional surveys in malaria-endemic communities
(Table III). Additionally, 70% of the published
studies were contributed by only three countries: Brazil, Venezuela and Colombia,
significantly limiting the geographical distribution of the available information.
Unfortunately,
not only are studies scarce but they are also not comparable because (i) the
designs of the studies were variable, (ii) the populations studied were different,
(iii) the definition of anaemia was not universal, (iv) it is difficult to thoroughly
understand the history of the infection, as several of the studies were cross-sectional
and not prospective, (v) co-infections were frequently present, such as bacteraemia
or helminth infection and (vi) patients with haemoglobinopathies or glucose-6-phosphate
dehydrogenase (G6PD) deficiencies presented at different prevalence rates. Therefore,
although severe anaemia was reported in these studies, it is not possible to
determine its prevalence, which appears to be low regardless. These findings
indicate the need for better-designed studies of malaria-related anaemia that
assess not only its overall prevalence, but also its association with malnutrition
and with common co-infections and diseases, such as helminth infections, bacterial
infections and hepatitis.
Clinical spectrum
of anaemia in human subjects - The clinical manifestations of malaria depend
on multiple factors from the parasite, parasite-host interactions and host factors,
including socio-economic conditions. Some of the host factors that have been
studied more in-depth include immunity, particularly the production of pro and
anti-inflammatory cytokines, genetic traits, α
or β-thalassemia,
Fy phenotype, sickle cell traits and age. Parasite factors such as endemicity,
drug resistance, Plasmodium species, parasite multiplication rates and
antigenic polymorphism are of great relevance for anaemia development. Moreover,
social-geographical factors, such as access to treatment, cultural and economic
factors, health policies and transmission intensity greatly contribute to increased
risk of anaemia.
Malarial anaemia
is usually normocytic and normochromic (Phillips et al. 1986, Bashawri et al.
2002), without spherocytes or schistocytes. However, the anaemia associated
with malaria can also be microcytic and hypochromic due to the high frequencies
of haemoglobinopathies and iron deficiency in endemic countries (Bashawri et
al. 2002).
Another key feature
of malarial anaemia is the presence of inadequate reticulocytosis, despite the
degree of anaemia. In acute uncomplicated malaria due to P. falciparum,
the haematocrit may be normal during the first 24 h after the onset of fever,
but afterwards there can be a progressive fall in the haematocrit level (Wickramasinghe
& Abdalla 2000) despite the initiation of anti-malarial treatment (Phillips
et al. 1986) and even in the absence of parasites in the blood smear (English
et al. 2002) or the administration of blood transfusions.
During malaria
infection, there are soluble derivatives released by the parasite that induce
bone marrow (BM) dysfunction. These derivatives are therefore implicated in
the pathogenesis of malarial anaemia (Silverman et al. 1987, Miller et al. 1989,
Jootar et al. 1993). This is reinforced by the observation that, as a consequence
of repeated infections or suboptimal treatment, children may be partially immunocompromised
and so asymptomatic during chronic P. falciparum infections. Despite
having very low Hb levels (Kurtzhals et al. 1999), these children display absolute
reticulocyte counts lower than expected for the degree of the anaemia (Wickramasinghe
& Abdalla 2000). These low counts might indicate some degree of BM dysfunction
(Kurtzhals et al. 1997). For P. vivax infection, it has been observed
that the decrease in Hb concentrations can be attributed to the activity of
the parasites. The destruction of erythrocytes is marked at the beginning of
the infection and erythrocytes do not return to pre-infection numbers in a short
period of time, despite the elimination of the infection. This can be primarily
explained by P. vivax invasion to reticulocytes, which prevents the establishment
of the normal erythrocyte population (Collins et al. 2003).
Immunopathology
- Although several mechanisms appear to participate in the generation of anaemia
in individuals acutely or chronically infected with malaria, the mechanisms
can nevertheless be grouped into two main categories: (i) destruction of cells
in the peripheral circulation and (ii) a reduction in or alteration of the production
of erythroid precursors. Although host genetics may exert some influence for
these two mechanisms, its role in anaemia pathogenesis is still poorly understood.
For instance, host genetic diversity may explain, to some extent, variations
in the frequency of anaemia when distinct areas are compared worldwide. The
Fy-negative genotype, which prevails on the Colombian Pacific coast, for example,
defines the higher prevalence of P. falciparum in that region, as compared
to other regions where P. vivax is more prevalent (Caicedo et al. 2009).
Likewise, there is evidence that FY*B/FY*X and FY*A/FY*X genotypes are associated
with low levels of P. vivax parasitism, which may favourably impact Hb
levels (Albuquerque et al. 2010). Recently, G6PD deficiency (Mediterranean type)
was shown to protect against P. vivax infection (Leslie et al. 2010);
however, protection by the African type, which is more prevalent in LA (Chagas
et al. 2009), has never been shown.
Peripheral destruction
of red blood cells (RBC) - An important part of the Plasmodium life
cycle occurs as an obligate intraerythrocytic parasite, where it differentiates
and multiplies at the expense of the host cell's nutrients up to the induction
of its burst. Each merozoite released either from the liver or from an erythrocyte
invades a new erythrocyte; in the case of P. vivax, its merozoites have
a preference for immature red cells (reticulocytes), whereas in the case of
P. falciparum, its merozoites invade erythrocytes of any age (Simpson et
al. 1999, Rayner et al. 2005). This parasite-specific invasion preference entails
the expression of receptors on the erythrocyte surfaces that are required for
an invasion. These include the Fy group antigens and reticulocyte-binding ligands
in the case of P. vivax (Galinski et al. 1992) or glycophorins A, B and
C, sialoglycoproteins expressed on human erythrocytes, in the case of P.
falciparum (JaŚkiewicz 2007). An obvious consequence of the parasite
multiplication and the periodic burst of schizonts is the rupture of infected
erythrocytes. Although this clearly contributes to the development of anaemia,
it does not appear to be sufficient to explain the levels of anaemia attained
in individuals exposed to the infection. It has been established that levels
of parasitaemia of > 50,000 parasites/µL are indicative of severe
falciparum malaria (WHO 2000). An infection level that corresponds to
the infection and destruction of approximately 1% of the total erythrocyte mass
could be easily replaced by erythropoiesis under normal conditions. However,
it seems that simultaneous to this mechanism, depuration of the parasitized
erythrocytes also occurs as a consequence of the phenomenon of erythrocyte rigidity.
This rigidity is induced by the transport of parasite antigens to the infected
erythrocyte membrane and is followed by the deformation of the membrane, opsonisation
by antibodies and complement and by macrophage activation (Wickramasinghe &
Abdalla 2000). However, in many cases, the severity of malaria anaemia does
not directly correlate with the degree of circulating parasitaemia (e.g. <
1%), though detected parasitaemia does not reflect the total parasite load,
as it does not take into account parasites sequestered in the microvasculature
(Nakazawa et al. 1995). Additionally, erythrocytes of malaria patients have
a decreased half-life compared to those of healthy individuals (Looareesuwan
et al. 1991). Moreover, epidemiological and mathematical models have shown that
between 8-12 non-parasitized erythrocytes may be destroyed for each Pf-RBC
parasitized (Jakeman et al. 1999, Price et al. 2001).
Although the precise
cause of the destruction of non-parasitized erythrocytes is unknown, several
mechanisms have been postulated: (i) the production of auto-antibodies against
the proteins that modify RBC membranes (Jakobsen et al. 1995), (ii) antibody-independent
phagocytosis of phosphatidyl-serine exposure, secondary to damage mediated by
reactive oxygen species (Bratosin et al. 1998, Serghides et al. 2003), (iii)
recognition of malarial antigens on infected erythrocytes by immunoglobulins
(Igs) and their further clearance by macrophages (Waitumbi et al. 2000), (iv)
complement-mediated phagocytosis and/or haemolysis (Ritter et al. 1993) and
(v) loss of complement regulatory proteins (CD35, CD55 and CD59) (Waitumbi et
al. 2000, Stoute et al. 2003). Furthermore, it has been documented that the
activities and absolute numbers of macrophages are increased during infection;
together with erythrocyte changes (decreased deformability and deposition of
Igs), this could assist in the depuration of non-parasitized erythrocytes during
infection (Mohan et al. 1995).
When dealing with
the poorly described P. vivax-induced anaemia phenomenon, another relevant
issue is the increasing body of evidence of P. vivax chloroquine resistance
in many endemic areas, including some regions of LA (Santana Filho et al. 2007),
which tends to increase peripheral parasitaemia for a longer period of time.
It has been demonstrated that in areas where chloroquine resistance has been
identified, severe disease (especially severe anaemia) is also frequent, which
raises the possibility of a causal effect (Price et al. 2009). Biomarkers of
resistance are urgently needed to validate this ecological association.
BM alterations
-The mechanisms of BM dysfunction induced by malaria appear to be multiple but
are still only partially known. The first observations of BM dysfunction during
malaria infection occurred over 60 years ago, when reticulocytopaenia was documented
in humans during P. falciparum and P. vivax infection. Afterwards,
it was shown that patients with acute P. falciparum infections had low
reticulocytaemia (or inappropriate reticulocytosis), which was accompanied with
suppression of erythropoiesis (erythroid hypoplasia) (Camacho et al. 1998).
Despite having increased cellularity in BM aspirates, there were no significant
differences in the total numbers of erythroblasts (Abdalla & Wickramasinghe
1998). These findings provided evidence of a decrease in the erythroid response
at the BM level. Children with chronic malaria, low parasitaemias (< 1%)
and severe anaemia display erythroid hyperplasia and dyserythropoiesis (Abdalla
et al. 1980, Wickramasinghe et al. 1989, Abdalla & Wickramasinghe 1998)
accompanied by ineffective erythropoiesis and decreases in circulating reticulocytes.
Inadequate production
of reticulocytes suggests insufficient erythropoiesis, which may be the result
of either hypoproliferative erythropoiesis or hyperproliferative but ineffective
erythropoiesis. Ineffective erythropoiesis is generally associated with intramedullar
destruction of erythroid precursors by erythrophagocytosis or dysplastic changes
of these precursors, which can be recognised by cytoplasmic vacuolation, abnormal
nucleus (bilobed), nuclear budding, formation of interchromatin and intracytoplasmatic
bridges and nuclear fragmentation.
It appears that
in acute and some chronic anaemia patients, there are two major forms of anaemia:
(i) anaemia with erythroid hypoplasia with or without dyserythropoiesis and
(ii) anaemia with erythroid hyperplasia and dyserythropoiesis. Acute malaria
infection in adults may be accompanied by a reduction in total erythropoietic
activity. In these cases, there may be normal BM or reduced cellularity with
erythroid hypoplasia. In cases of high parasitaemia, there may even be ineffective
erythropoiesis of the residual erythropoietic activity (hypoproliferative erythropoiesis).
Several studies
have documented a loss of precursor cells in the BM (Dörmer et al. 1983),
reduced use of iron by the erythrocytes and dysplasic changes in the BM (Knuttgen
1987). However, in other studies, no evidence of dyserythropoiesis was found
in children with acute P. falciparum infection (Das et al. 1999). Therefore,
it has been proposed that, in contrast to chronic malaria, dyserythropoiesis
plays a minor role in the pathogenesis of anaemia during an acute malaria infection
(Das et al. 1999, Jakeman et al. 1999). Some studies suggest that ineffective
erythropoiesis and dyserythropoiesis play greater roles (Abdalla et al. 1980,
Weatherall et al. 1983) in chronic malaria infection, which may be accompanied
by a severe anaemia characterised by an erythroid hyperplasia and dysplasia,
some degree of erythrophagocytosis and low levels of reticulocytes. In this
case, the dyserythropoiesis is directly related to malaria and is not caused
by deficiencies of folate, vitamin B12 or iron (Abdalla et al. 1984).
However, ineffective
erythropoiesis during malaria could develop through different mechanisms, such
as altered Hb synthesis in vitro and premature death of normoblasts (Srichaikul
et al. 1973, 1976).
Susceptible
populations - In malaria-endemic regions, the most susceptible populations
to suffer severe and complicated disease, including anaemia, are children <
five years of age and pregnant women. Although older children and adults still
suffer repeated malaria infections, the disease frequency and severity is progressively
reduced. This clinical immunity does not develop in areas of low endemicity
or seasonal exposure to parasites; therefore, the disease affects all groups
in these regions (Miller et al. 1994). Pregnant women, although previously clinically
immune, become more susceptible to developing the pathogenic processes that
affect both the mother and the foetus, and subsequently the newborn.
Two of the most
feared malaria complications that are associated with an increased mortality,
especially in children and pregnant women, are cerebral malaria and severe anaemia,
with mortality rates of 5.6-16% in children (Marsh et al. 1995) and approximately
6% in pregnant women (Granja et al. 1998, Menendez et al. 2000, Weatherall et
al. 2002). As mentioned, immunological factors and mechanisms appear to have
great relevance in the anaemia pathogenesis during a malaria infection. Beside
the role of the antibodies that are specific to malarial antigens and are exported
to erythrocyte membranes, the potential role of the erythrocyte-targeted auto-antibodies
and that of complement activation and cytokine imbalances are associated with
an increased anaemia severity in children with malaria. Pro-inflammatory cytokines,
such as tumour necrosis factor-alpha (TNF-α)
and interleukin (IL)-6, are elevated during an acute malarial infection (Kern
et al. 1989, Lyke et al. 2004) while anti-inflammatory cytokines, such as IL-10,
are substantially decreased (Kurtzhals et al. 1998, 1999, Akanmori et al. 2000).
Furthermore, elevated TNF-α
levels are associated with an increased anaemia severity in children with malaria
(Shaffer et al. 1991) and a low IL-10/TNF-α
ratio is associated with an enhanced anaemia severity, suggesting that the relative
expression of cytokines in the inflammatory milieu is an important determinant
of severe malarial anaemia (Othoro et al. 1999, Perkins et al. 2000). It has
also been observed that increased levels of TNF-α
are associated with a decrease in erythroid progenitor cells, decreased iron
uptake by erythrocytes, erythrophagocytosis of nucleated erythroblasts and dyserythropoiesis
(Phillips et al. 1986, Silverman et al. 1987, Clark & Chaudhri 1988, Miller
et al. 1989, Taverne et al. 1994). Moreover, recent studies show that hepcidin,
a 25-amino-acid protein produced in the liver, is associated with the anaemia
of inflammation in humans, where its production is increased 100-fold, resulting
in both an impaired iron uptake in the gut and iron sequestration in macrophages
(Ganz 2003, Means 2004).
Between 60-80%
of Hb is degraded during the intraerythrocytic cycle, releasing haemozoin (Hz)
and amino acids, which are used by the parasite to produce proteins. The presence
of Hz in the cytoplasm of polymorphonuclear leukocytes and monocytes appears
to be associated with the severity of the malarial infection, as it seems that
cytoplasmic Hz is more frequently found in the complicated malaria cases than
in the uncomplicated cases (Nguyen et al. 1995, Amodu et al. 1998, Lyke et al.
2003, López et al. 2004).
Experimental
animal models of malarial anaemia - In-depth haematological studies in humans
with malarial anaemia pose a number of ethical and technical challenges that
preclude invasive procedures, particularly BM analyses over the course of the
infection. The development of experimental animal models is therefore critical
for understanding the mechanisms involved in the pathogenesis of severe anaemia.
Although some molecular bases of malarial anaemia could be shared by several
Plasmodium species, experimental evidence suggests that species-specific
factors play significant roles. For example, a larger proportion of the non-infected
RBCs are removed by erythrophagocytosis in P. vivax-infected individuals
- it is estimated that ~32 non-infected RBCs are destroyed per every P. vivax-infected
RBC (Collins et al. 2003), compared to approximately eight non-infected RBCs
destroyed per every P. falciparum-infected RBC (Jakeman et al. 1999).
Four rodent malaria
parasite species (Plasmodium berghei, Plasmodium chabaudi, Plasmodium
vinckei and Plasmodium yoelii), have been extensively used to study
malaria pathogenesis, including anaemia, due to their distinctive erythrocyte
invasion profiles, which are similar to those observed with human parasites
(Lamb et al. 2006). However, several features, such as anaemia in the presence
of hyperparasitaemia and extramedullar erythropoiesis, which are frequently
observed in rodents, are rare events in human malarial infections (Silverman
et al. 1987, Yap & Stevenson 1992). Anaemia research in rodent models allows
immunological studies and manipulations that are more difficult in other animal
models, such as primates. Examples include in vivo depletion of macrophages
and CD4+ T cells, comparisons of resistant vs. susceptible strains,
experiments involving cytokine knockout mice, such as IL-10 (Linke et al. 1996)
and macrophage migration inhibitory factor-deficient mice (Stevenson et al.
2001, McDevitt et al. 2006). Non-human primate models appear to be more relevant
in LA, due to their abundance in the region and the availability of several
primate colonies.
New World monkeys
(Aotus and Saimiri) have been used extensively for vaccine trials
and drug testing using P. falciparum and P. vivax-adapted strains
(Collins 1992, Obaldia 2001, Herrera et al. 2002). Malaria semi-immune Aotus
monkeys immunised with merozoite vaccine candidates or exposed to P. falciparum
were protected from hyperparasitaemia, but were more likely to develop severe
anaemia after a second challenge (Egan et al. 2002, Jones et al. 2002). In this
experimental model, low Hb levels were associated with low reticulocyte counts,
suggesting that the ineffective erythropoiesis and removal of non-infected erythrocytes
are at least part of the aetiological factors involved (Egan et al. 2002). Aotus
monkeys that self-control parasite patency or that received anti-malaria treatment
exhibited a robust reticulocytosis, indicating a direct effect of the parasite
on erythroid progenitors. Interestingly, immunisation of Aotus monkeys
with the P. falciparum CIDR1α
domain of PfEMP1, a protein involved in sequestration, prevents the development
of anaemia after re-infection (Makobongo et al. 2006). Aotus also appears
to be a good model to study the role of hepcidin homeostasis. Although little
is known about the relationship between hepcidin and malarial anaemia, one study
developed in Colombia with the Aotus model showed that hepcidin levels
decreased throughout the experiment in malaria and mock-infected animals (Llanos
2008). Regardless of this finding, it remains an area for further research.
Methodologies have
been implemented to study the BM compartment in Aotus monkeys experimentally
infected with P. falciparum and P. vivax (Llanos et al. 2006).
Interestingly, on-going studies have confirmed that high numbers of normoblasts
are present in BM aspirates, suggesting that erythropoiesis is effective (unpublished
data) (Llanos 2008). However, the molecular mechanisms involved in severe anaemia
in semi-immune Aotus monkeys remain unknown (Egan et al. 2002).
Simian malaria
parasites have been a critical resource for understanding the biology of Plasmodium
(Brown & Brown 1965, Coatney 1968) for facilitating the development of anti-malarial
drugs (Omar et al. 1973) and for characterising the mechanisms involved in the
physiopathology of severe malaria (Davison et al. 1998). Unfortunately, comprehensive
investigations on malaria pathogenesis have not been addressed using these experimental
models (Galinski & Barnwell 2008). Plasmodium coatneyi and Plasmodium
fragile share with P. falciparum the ultrastructural features that
are involved in parasite sequestration and rosetting. Therefore, these parasite
species mimic clinical complications of cerebral malaria associated with P.
falciparum infections in humans (Kawai et al. 1993, Fujioka et al. 1994).
P. coatneyi infection in macaques has also been associated with placental
malaria, thrombocytopaenia, anaemia and disseminated intravascular coagulation
(Kawai et al. 1993, Sein et al. 1993, Nakano et al. 1996, Smith et al. 1996,
Davison et al. 1998, Collins et al. 2001, Moreno et al. 2007). Plasmodium
cynomolgi is phylogenetically related to P. vivax and shares its
capacity to produce hypnozoites (Krotoski et al. 1982). The patterns of relapses
described in P. vivax are also present in rhesus macaques exposed to
P. cynomolgi sporozoites, making this model ideal for studying the mechanisms
of chronic anaemia (Schmidt 1986). Comparative experiments using P. coatneyi
and P. cynomolgi in rhesus macaques are required to define whether the
molecular mechanisms underlying the pathogenesis of malarial anaemia are shared
between these two species. Unfortunately, such studies are restricted to primate
centres outside LA.
Conclusions
and perspectives - Beside the acknowledged importance of anaemia as a cause
of morbidity and mortality in Africa and other endemic regions, little is known
about its prevalence and burden in the malaria-endemic regions of LA. The few
studies reported from this region appear to indicate that the incidence of severe
anaemia is significantly lower than that reported from Africa or Asia. However,
considering that specific haematological changes associated with malaria infection
may vary with the level of malaria endemicity (Idro et al. 2006), nutritional
status (Friedman et al. 2005), demographic factors (Barcus et al. 2007), malaria
immunity (Langhorne et al. 2008) and the parasite species, it is essential to
study and characterise the epidemiology and mechanisms involved in malarial
anaemia in endemic regions of LA. The National Institutes of Health-National
Institute of Allergy and Infectious Diseases is currently funding the establishment
of the Centro Latino Americano de Investigación en Malaria (CLAIM) as
a Centre of Excellence for Malaria Research (ICEMR). The Centre aims, as a priority,
to determine the prevalence and severity of haematological manifestations attributable
to malaria infection and their association with concomitant immune status, including
nutritional factors and hel-minth coinfection. Although studies would initially
cover only Colombia, Panama, Peru and Guatemala, further expansion of malaria
research, including anaemia, would be extended to other countries of LA, to
the Caribbean region and Brazil. CLAIM already interacts with the Brazilian
Malaria Network, which aims to describe the determinants of P. vivax-related
severe anaemia in hospitalised patients from a tertiary care institution, specifically
focussing on the impacts of host genetics (G6PD deficiency and Fy genotypes)
and chloroquine resistance.
These multi-country
regional projects will allow the comparison of pathologies in settings with
different malaria transmission intensities in communities with great ethnic,
occupational and immune diversities. Moreover, the availability of the Aotus
monkey animal model in several countries of the LA region represents a valuable
resource to address important questions regarding malaria pathogenesis that
cannot be studied in the human populations. Indeed, the accessibility to BM
aspirates of non-human primates vaccinated with human malaria vaccine candidates
opens an interesting new area of research to complement the analyses of the
influences of specific antimalarial immune responses in the generation of anaemia.
ACKNOWLEDGEMENTS
To the participation
of endemic communities from malaria endemic areas of Colombia and Brazil, who
kindly accepted to participate in part of the studies mentioned here, and to
Paola Rojas, for her technical support to prepare this paper.
References
- Abdalla S, Weatherall
DJ, Wickramasinghe SN, Hughes M 1980. The anaemia of P. falciparum malaria.
Br J Haematol 46: 171-183.
- Abdalla SH, Wickramasinghe
SN 1998. A study of erythroid progenitor cells in the bone marrow of Gambian
children with falciparum malaria. Clin Lab Haematol 10: 33-40.
- Abdalla SH, Wickramasinghe
SN, Weatherall DJ 1984. The deoxyuridine suppression test in severe anaemia
following Plasmodium falciparum malaria. Trans R Soc Trop Med Hyg
78: 60-63.
- Akanmori BD, Kurtzhals
JA, Goka BQ, Adabayeri V, Ofori MF, Nkrumah FK, Behr C, Hviid L 2000. Distinct
patterns of cytokine regulation in discrete clinical forms of Plasmodium
falciparum malaria. Eur Cytokine Netw 11: 113-118.
- Akhwale WS, Lum
JK, Kaneko A, Eto H, Obonyo C, Björkman A, Kobayakawa T 2004. Anemia and
malaria at different altitudes in the western highlands of Kenya. Acta Trop
91: 167-175.
- Albuquerque SR,
Cavalcante F de O, Sanguino EC, Tezza L, Chacon F, Castilho L, dos Santos MC
2010. FY polymorphisms and vivax malaria in inhabitants of Amazonas State, Brazil. Parasitol Res 106: 1049-1053.
- Alexandre MA, Ferreira
CO, Siqueira AM, Magalhaes BL, Mourao MP, Lacerda MV, Alecrim MG 2010. Severe Plasmodium vivax malaria, Brazilian Amazon. Emerg Infect Dis 16:
1611-1614.
- Amaral CNd, Albuquerque
YDd, Pinto AYDN, Souza JMD 2003. A importancia do perfil clínico-laboratorial
no diagnostico diferencial entre malaria e hepatite aguda viral. J Pediatr 79: 429-434.
- Amodu OK, Adeyemo
AA, Olumese PE, Gbadegesin RA 1998. Intraleucocytic malaria pigment and clinical
severity of malaria in children. Trans R Soc Trop Med Hyg 92: 54-56.
- Andrade BB, Reis-Filho
A, Souza-Neto SM, Clarencio J, Camargo LM, Barral A, Barral-Netto M 2010. Severe Plasmodium vivax malaria exhibits marked inflammatory imbalance. Malar
J 9: 13.
- Bacon DJ, McCollum
AM, Griffing SM, Salas C, Soberon V, Santolalla M, Haley R, Tsukayama P, Lucas
C, Escalante AA, Udhayakumar V 2009. Dynamics of malaria drug resistance patterns
in the Amazon Basin region following changes in Peruvian national treatment
policy for uncomplicated malaria. Antimicrob Agents Chemother 53: 2042-2051.
- Barcus MJ, Basri
H, Picarima H, Manyakori C, Sekartuti, Elyazar I, Bangs MJ, Maguire JD, Baird
JK 2007. Demographic risk factors for severe and fatal vivax and falciparum malaria among hospital admissions in northeastern Indonesian Papua. Am J
Trop Med Hyg 77: 984-991.
- Bardales Tuesta
F, De la Puente-Olortegui C 2000. Manejo en UCI de la malaria falciparum severa y complicada. Bol Soc Peru Med Interna 13: 30-39.
- Barraviera B, Machado
PE, Meira DA 1988. Glutathione reductase activity and its relation with riboflavin
levels measured by methemoglobin reduction by cystamine in patients with malaria
(preliminary report). Rev Inst Med Trop Sao Paulo 30: 107-108.
- Bashawri LA, Mandil
AA, Bahnassy AA, Ahmed MA 2002. Malaria: hematological aspects. Ann Saudi
Med 22: 372-376.
- Blair S, Alvares
G, Campuzano G 1997. Relación entre anemia y malaria en una población
rural de Colombia. Boletín de la direccion de malariologia y saneamiento
ambiental 37: 7-12.
- Bratosin D, Mazurier
J, Tissier JP, Estaquier J, Huart JJ, Ameisen JC, Aminoff D, Montreuil J 1998.
Cellular and molecular mechanisms of senescent erythrocyte phagocytosis by macrophages.
A review. Biochimie 80: 173-195.
- Brown KN, Brown
IN 1965. Immunity to malaria: antigenic variation in chronic infections of Plasmodium
knowlesi. Nature 208: 1286-1288.
- Caicedo O, Ramirez
O, Mourao MP, Ziadec J, Perez P, Santos JB, Quinones F, Alecrim MG, Arevalo-Herrera
M, Lacerda MV, Herrera S 2009. Comparative hematologic analysis of uncomplicated
malaria in uniquely different regions of unstable transmission in Brazil and
Colombia. Am J Trop Med Hyg 80: 146-151.
- Caicedo O, Villamor
E, Forero Y, Ziade J, Pérez P, Quiñones F, Arévalo-Herrera
M, Herrera S 2010. Relation between vitamin B12 and folate status and hemoglobin
concentration and parasitemia during acute malaria infections in Colombia. Acta
Tropica 114: 17-21.
- Camacho LH, Gordeuk
VR, Wilairatana P, Pootrakul P, Brittenham GM, Looareesuwan S 1998. The course
of anaemia after the treatment of acute, falciparum malaria. Ann Trop Med
Parasitol 92: 525-537.
- Cardoso MA, Ferreira
MU, Camargo LM, Szarfarc SC 1992. Anemia in a population from an endemic area
of malaria, Rondonia (Brazil). Rev Saude Publica 26: 161-166.
- Cardoso MA, Ferreira
MU, Camargo LM, Szarfarc SC 1994. Anaemia, iron deficiency and malaria in a
rural community in Brazilian Amazon. Eur J Clin Nutr 48: 326-332.
- Carmona-Fonseca
2008. Vivax malaria in children: clinical features and response to chloroquine. Colomb Med 39: 364-377.
- Castano AT, Pineros
Jimenez JG, Trujillo SB, Carmona-Fonseca J 2006. Clinica de la malaria complicada
debida a P. falciparum. Estudio de casos y controles en Tumaco y Turbo
(Colombia). Iatreia 19: 339-355.
- Chagas EC, Nascimento
CT, Santana Filho FS, Botto-Menezes CH, Martinez-Espinosa FE 2009. Impact of
malaria during pregnancy in the Amazon Region. Rev Panam Salud Publica 26:
203-208.
- Chiriboga J, Machiavello
S, Luglio M, Aiscorbe R 1950. Malaria, uncinariasis, nutricion y anemia en los
tropicos. Medicina 10: 353-366.
- Clark IA, Chaudhri
G 1988. Tumour necrosis factor may contribute to the anaemia of malaria by causing
dyserythropoiesis and erythrophagocytosis. Br J Haematol 70: 99-103.
- Coatney GR 1968.
Simian malarias in man: facts, implications and predictions. Am J Trop Med
Hyg 17: 147-155.
- Collins WE 1992.
South American monkeys in the development and testing of malarial vaccines -
A review. Mem Inst Oswaldo Cruz 87 (Suppl. III): 401-406.
- Collins WE, Jeffery
GM, Roberts JM 2003. A retrospective examination of anemia during infection
of humans with Plasmodium vivax. Am J Trop Med Hyg 68: 410-412.
- Collins WE, Warren
M, Sullivan JS, Galland GG 2001. Plasmodium coatneyi: observations on
periodicity, mosquito infection and transmission to Macaca mulatta monkeys. Am J Trop Med Hyg 64: 101-110.
- Corredor V, Murillo
C, Echeverry DF, Benavides J, Pearce RJ, Roper C, Guerra AP, Osorio L 2010.
Origin and dissemination across the Colombian Andes mountain range of sulfadoxine-pyrimethamine
resistance in Plasmodium falciparum. Antimicrob Agents Chemother 54:
3121-3125.
- Cortese JF, Caraballo
A, Contreras CE, Plowe CV 2002. Origin and dissemination of Plasmodium falciparum drug-resistance mutations in South America. J Infect Dis 186: 999-1006.
- Das BS, Nanda NK,
Rath PK, Satapathy RN, Das DB 1999. Anaemia in acute, Plasmodium falciparum malaria in children from Orissa state, India. Ann Trop Med Parasitol 93:
109-118.
- Davison BB, Cogswell
FB, Baskin GB, Falkenstein KP, Henson EW, Tarantal AF, Krogstad DJ 1998. Plasmodium
coatneyi in the rhesus monkey (Macaca mulatta) as a model of malaria
in pregnancy. Am J Trop Med Hyg 59: 189-201.
- De Souza JM 1983.
A Phase II clinical trial of mefloquine in Brazilian male subjects. Bull
World Health Organ 61: 815-820.
- Dicko A, Mantel
C, Thera MA, Doumbia S, Diallo M, Diakite M, Sagara I, Doumbo OK 2003. Risk
factors for malaria infection and anemia for pregnant women in the Sahel area
of Bandiagara, Mali. Acta Trop 89: 17-23.
- Dörmer P,
Dietrich M, Kern P, Horstmann RD 1983. Ineffective erythropoiesis in acute human P. falciparum malaria. Blut 46: 279-288.
- Echeverri M, Tobón
A, Alvarez G, Carmona J, Blair S 2003. Clinical and laboratory findings of Plasmodium
vivax malaria in Colombia, 2001. Rev Inst Med Trop Sao Paulo 45:
29-34.
- Egan AF, Fabucci
ME, Saul A, Kaslow DC, Miller LH 2002. Aotus New World monkeys: model
for studying malaria-induced anemia. Blood 99: 3863-3866.
- English M, Ahmed
M, Ngando C, Berkley J, Ross A 2002. Blood transfusion for severe anaemia in
children in a Kenyan hospital. Lancet 359: 494-495.
- Espinoza E, Hidalgo
L, Chedraui P 2005. The effect of malarial infection on maternal-fetal outcome
in Ecuador. J Matern Fetal Neonatal Med 18: 101-105.
- Faich GA, Mason
J 1975. The prevalence and relationships of malaria, anemia and malnutrition
in a coastal area of El Salvador. Am J Trop Med Hyg 24: 161-167.
- Fernandes AAM,
Carvalho LJM, Zanini GM, Ventura AMRS, Souza JM, Cotias PM, Silva-Filho IL,
Daniel-Ribeiro CT 2008. Similar cytokine responses and degrees of anemia in
patients with Plasmodium falciparum and Plasmodium vivax infections
in the Brazilian Amazon Region. Clin Vaccine Immunol 15: 650-658.
- Fernandez RD, Garcia
Y, Alger J 2001. Malaria y embarazo: observaciones clinico-epidemiologicas en
dos zonas geograficas de Honduras. Rev Med Hondur 69: 8-18.
- Ferreira MU, da
Silva-Nunes M, Bertolino CN, Malafronte RS, Muniz PT, Cardoso MA 2007. Anemia
and iron deficiency in school children, adolescents and adults: a community-based
study in rural Amazonia. Am J Public Health 97: 237-239.
- Friedman JF, Kwena
AM, Mirel LB, Kariuki SK, Terlouw DJ, Phillips-Howard PA, Hawley WA, Nahlen
BL, Shi YP, ter Kuile FO 2005. Malaria and nutritional status among pre-school
children: results from cross-sectional surveys in western Kenya. Am J Trop
Med Hyg 73: 698-704.
- Fujioka H, Millet
P, Maeno Y, Nakazawa S, Ito Y, Howard RJ, Collins WE, Aikawa M 1994. A nonhuman
primate model for human cerebral malaria: rhesus monkeys experimentally infected
with Plasmodium fragile. Exp Parasitol 78: 371-376.
- Galinski M, Barnwell
J 2008. Plasmodium vivax: who cares? Malaria J 7: S9.
- Galinski MR, Medina
CC, Ingravallo P, Barnwell JW 1992. A reticulocyte-binding protein complex of Plasmodium vivax merozoites. Cell 69: 1213-1226.
- Ganz T 2003. Hepcidin,
a key regulator of iron metabolism and mediator of anemia of inflammation. Blood
102: 783-788.
- García-Casal
MN, Leets I, Bracho C, Hidalgo M, Bastidas G, Gomez A, Peña A, Pérez
H 2008. Prevalence of anemia and deficiencies of iron, folic acid and vitamin
B12 in an indigenous community from the Venezuelan Amazon with a high incidence
of malaria. Arch Latinoam Nutr 58: 12-18.
- Genton B, D'Acremont
V, Rare L, Baea K, Reeder JC, Alpers MP, Muller I 2008. Plasmodium vivax and mixed infections are associated with severe malaria in children: a prospective
cohort study from Papua New Guinea. PLoS Med 5: e127.
- Gomez E, Lopez
E, Ache A 2009. Malaria y embarazo. Parroquia San Isidro, municipio Sifontes,
estado Bolivar, Venezuela, anos 2005-2006. Invest Clin 50: 455-464.
- González
L, Guzmán M, Carmona-Fonseca J, Lopera T, Blair S 2000. Características
clínicoepidemiológicas de 291 pacientes hospitalizados por malaria
en Medellín (Colombia). Acta Med Col 25: 163-170.
- Granja AC, Machungo
F, Gomes A, Bergström S, Brabin B 1998. Malaria-related maternal morbidity
in urban Mozambique. Ann Trop Med Parasitol 92: 257-263.
- Grenfell P, Fanello
CI, Magris M, Goncalves J, Metzger WG, Vivas-Martínez S, Curtis C, Vivas
L 2008. Anaemia and malaria in Yanomami communities with differing access to
healthcare. Trans R Soc Trop Med Hyg 102: 645-652.
- Guerra CA, Gikandi
PW, Tatem AJ, Noor AM, Smith DL, Hay SI, Snow RW 2008. The limits and intensity
of Plasmodium falciparum transmission: implications for malaria control
and elimination worldwide. PLoS Med 5: e38.
- Guerra CA, Howes
RE, Patil AP, Gething PW, Van Boeckel TP, Temperley WH, Kabaria CW, Tatem AJ,
Manh BH, Elyazar IRF, Baird JK, Snow RW, Hay SI 2010. The international limits
and population at risk of Plasmodium vivax transmission in 2009. PLoS
Negl Trop Dis 4: e774.
- Gusmao R 1999.
Overview of malaria control in the Americas. Parassitologia 41: 355-360.
- Gutierrez M, Maldonado
I, Balliache N, Contreras I 1998. Anticuerpos antifosfolipidos en malaria. Med
Interna (Caracas) 14: 142-152.
- Hadley TJ, Peiper
SC 1997. From malaria to chemokine receptor: the emerging physiologic role of
the Duffy blood group antigen. Blood 89: 3077-3091.
- Hamer DH, MacLeod
WB, Addo-Yobo E, Duggan CP, Estrella B, Fawzi WW, Konde-Lule JK, Mwanakasale
V, Premji ZG, Sempertegui F, Ssengooba FP, Yeboah-Antwi K, Simon JL 2003. Age,
temperature and parasitaemia predict chloroquine treatment failure and anaemia
in children with uncomplicated Plasmodium falciparum malaria. Trans
R Soc Trop Med Hyg 9: 422-428.
- Herrera S, Perlaza
BL, Bonelo A, Arevalo-Herrera M 2002. Aotus monkeys: their great value
for anti-malaria vaccines and drug testing. Int J Parasitol 32: 1625-1635.
- Idro R, Aloyo J,
Mayende L, Bitarakwate E, John CC, Kivumbi GW 2006. Severe malaria in children
in areas with low, moderate and high transmission intensity in Uganda. Trop
Med Int Health 11: 115-124.
- Jakeman GN, Saul
A, Hogarth WL, Collins WE 1999. Anaemia of acute malaria infections in non-immune
patients primarily results from destruction of uninfected erythrocytes. Parasitology
119: 127-133.
- Jakobsen PH, Bate
CA, Taverne J, Playfair JH 1995. Malaria: toxins, cytokines and disease. Parasite
Immunol 17: 223-231.
- Jarude R, Trindade
R, Tavares-Neto J 2003. Malária em grávidas de uma maternidade
pública de Rio Branco (Acre, Brasil). Rev Bras Ginecol Obstet 25:
149-154.
- JaŚkiewicz
E 2007. Glycophorins of human erythrocytes as receptors for the malaria parasite Plasmodium falciparum. Postepy Hig Med Dosw 61: 718-724.
- Jones TR, Stroncek
DF, Gozalo AS, Obaldia N 3rd, Andersen EM, Lucas C, Narum DL, Magill AJ, Sim
BK, Hoffman SL 2002. Anemia in parasite- and recombinant protein-immunized Aotus monkeys infected with Plasmodium falciparum. Am J Trop Med Hyg 66:
672-679.
- Jootar S, Chaisiripoomkere
W, Pholvicha P, Leelasiri A, Prayoonwiwat W, Mongkonsvitragoon W, Srichaikul
T 1993. Suppression of erythroid progenitor cells during malarial infection
in Thai adults caused by serum inhibitor. Clin Lab Haematol 15: 87-92.
- Kawai S, Aikawa
M, Kano S, Suzuki M 1993. A primate model for severe human malaria with cerebral
involvement: Plasmodium coatneyi-infected Macaca fuscata. Am
J Trop Med Hyg 48: 630-636.
- Kern P, Hemmer
CJ, Van Damme J, Gruss HJ, Dietrich M 1989. Elevated tumor necrosis factor alpha
and interleukin-6 serum levels as markers for complicated Plasmodium falciparum malaria. Am J Med 87: 139-143.
- Knuttgen H 1987.
The bone marrow of non-immune Europeans in acute malaria infection: a topical
review. Ann Trop Med Parasitol 81: 567-576.
- Kochar DK, Das
A, Kochar SK, Saxena V, Sirohi P, Garg S, Kochar A, Khatri MP, Gupta V 2009.
Severe Plasmodium vivax malaria: a report on serial cases from Bikaner
in northwestern India. Am J Trop Med Hyg 80: 194-198.
- Krotoski WA, Garnham
PC, Bray RS, Krotoski DM, Killick-Kendrick R, Draper CC, Targett GA, Guy MW
1982. Observations on early and late post-sporozoite tissue stages in primate
malaria. I. Discovery of a new latent form of Plasmodium cynomolgi (the
hypnozoite) and failure to detect hepatic forms within the first 24 hours after
infection. Am J Trop Med Hyg 31: 24-35.
- Kurtzhals JA, Adabayeri
V, Goka BQ, Akanmori BD, Oliver-Commey JO, Nkrumah FK, Behr C, Hviid L 1998.
Low plasma concentrations of interleukin-10 in severe malarial anaemia compared
with cerebral and uncomplicated malaria. Lancet 351: 1768-1772.
- Kurtzhals JA, Akanmori
BD, Goka BQ, Adabayeri V, Nkrumah FK, Behr C, Hviid L 1999. The cytokine balance
in severe malarial anemia. J Infect Dis 180: 1753-1755.
- Kurtzhals JA, Rodrigues
O, Addae M, Commey JO, Nkrumah FK, Hviid L 1997. Reversible suppression of bone
marrow response to erythropoietin in Plasmodium falciparum malaria. Br
J Haematol 97: 169-174.
- Lamb TJ, Brown
DE, Potocnik AJ, Langhorne J 2006. Insights into the immunopathogenesis of malaria
using mouse models. Expert Rev Mol Med 8: 1-22.
- Langhorne J, Ndungu
FM, Sponaas AM, Marsh K 2008. Immunity to malaria: more questions than answers. Nat Immunol 9: 725-732.
- Leslie T, Briceno
M, Mayan I, Mohammed N, Klinkenberg E, Sibley CH, Whitty CJ, Rowland M 2010.
The impact of phenotypic and genotypic G6PD deficiency on risk of Plasmodium
vivax infection: a case-control study amongst Afghan refugees in Pakistan. PLoS Med 7: e1000283.
- Linke A, Kühn
R, Müller W, Honarvar N, Li C, Langhorne J 1996. Plasmodium chabaudi
chabaudi: differential susceptibility of gene-targeted mice deficient in
IL-10 to an erythrocytic-stage infection. Exp Parasitol 84: 253-263.
- Lisker R, Loria
A, Cordova MS 1965. Studies on several genetic hematological traits of the Mexican
population. 8. Hemoglobin S, glucose-6-phosphate dehydrogenase deficiency and
other characteristics in a malarial region. Am J Hum Genet 17: 179-187.
- Llanos C 2008.
Plasmodium falciparum-induced anemia and dyserythropoiesis in naÏve
Aotus monkeys, Valle University, 95 pp.
- Llanos C, Quintero
G, Castellanos A, Arevalo-Herrera M, Herrera S 2006. Surgical bone marrow aspiration
in Aotus lemurinus griseimembra. J Med Primatol 35: 131-135.
- Looareesuwan S,
Davis TM, Pukrittayakamee S, Supanaranond W, Desakorn V, Silamut K, Krishna
S, Boonamrung S, White NJ 1991. Erythrocyte survival in severe falciparum malaria. Acta Trop 48: 263-270.
- López ML,
Arango EM, Arias LR, Carmona-Fonseca J 2004. Hemozoína intraleucocitaria
como indicador de malaria complicada por Plasmodium falciparum. Acta
Med Colomb 29: 80-87.
- Lyke KE, Burges
R, Cissoko Y, Sangare L, Dao M, Diarra I, Kone A, Harley R, Plowe CV, Doumbo
OK, Sztein MB 2004. Serum levels of the proinflammatory cytokines interleukin-1
beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha and IL-12(p70)
in Malian children with severe Plasmodium falciparum malaria and matched
uncomplicated malaria or healthy controls. Infect Immun 72: 5630-5637.
- Lyke KE, Diallo
DA, Dicko A, Kone A, Coulibaly D, Guindo A, Cissoko Y, Sangare L, Coulibaly
S, Dakouo B, Taylor TE, Doumbo OK, Plowe CV 2003. Association of intraleukocytic Plasmodium falciparum malaria pigment with disease severity, clinical
manifestations and prognosis in severe malaria. Am J Trop Med Hyg 69:
253-259.
- Magris M, Rubio-Palis
Y, Alexander N, Ruiz B, Galván N, Frias D, Blanco M, Lines J 2007. Community-randomized
trial of lambdacyhalothrin-treated hammock nets for malaria control in Yanomami
communities in the Amazon Region of Venezuela. Trop Med Int Health 12:
392-403.
- Makobongo MO, Keegan
B, Long CA, Miller LH 2006. Immunization of Aotus monkeys with recombinant
cysteine-rich interdomain region 1 alpha protects against severe disease during Plasmodium falciparum reinfection. J Infect Dis 193: 731-740.
- Marquino W, Ylquimiche
L, Hermenegildo Y, Palacios AM, Falconi E, Cabezas C, Arrospide N, Gutierrez
S, Ruebush ITK 2005. Efficacy and tolerability of artesunate plus sulfadoxine-pyrimethamine
and sulfadoxine-pyrimethamine alone for the treatment of uncomplicated Plasmodium
falciparum malaria in Peru. Am J Trop Med Hyg 72: 568-572.
- Marsh K, Forster
D, Waruiru K, Mwangi I, Winstanley M, Marsh V, Newton C, Winstanley P, Warn
P, Peshu N, Pasvol G 1995. Indicators of life-threatening malaria in African
children. N Engl J Med 332: 1399-1404.
- McCollum AM, Mueller
K, Villegas L, Udhayakumar V, Escalante AA 2007. Common origin and fixation
of Plasmodium falciparum dhfr and dhps mutations associated with sulfadoxine-pyrimethamine
resistance in a low-transmission area in South America. Antimicrob Agents
Chemother 51: 2085-2091.
- McDevitt MA, Xie
J, Shanmugasundaram G, Griffith J, Liu A, McDonald C, Thuma P, Gordeuk VR, Metz
CN, Mitchell R, Keefer J, David J, Leng L, Bucala R 2006. A critical role for
the host mediator macrophage migration inhibitory factor in the pathogenesis
of malarial anemia. J Exp Med 203: 1185-1196.
- Means RT Jr. 2004.
Hepcidin and anaemia. Blood Rev 18: 219-225.
- Melo GC, Reyes-Lecca
RC, Vitor-Silva S, Monteiro WM, Martins M, Benzecry SG, Alecrim MG, Lacerda
MV 2010. Concurrent helminthic infection protects schoolchildren with Plasmodium
vivax from anemia. PLoS ONE 5: e11206.
- Menendez C, Fleming
AF, Alonso PL 2000. Malaria-related anaemia. Parasitol Today 16: 469-476.
- Menendez-Capote
R, Diaz Perez L, Luzardo Suarez C 1997. Hemolisis y tratamiento con primaquina.
Informe preliminar. Rev Cubana Med Trop 49: 136-138.
- Miller KL, Silverman
PH, Kullgren B, Mahlmann LJ 1989. Tumor necrosis factor alpha and the anemia
associated with murine malaria. Infect Immun 57: 1542-1546.
- Miller LH, Good
MF, Milon G 1994. Malaria pathogenesis. Science 264: 1878-1883.
- Mohan K, Dubey
ML, Ganguly NK, Mahajan RC 1995. Plasmodium falciparum: role of activated
blood monocytes in erythrocyte membrane damage and red cell loss during malaria. Exp Parasitol 80: 54-63.
- Moreno A, García
A, Cabrera-Mora M, Strobert E, Galinski MR 2007. Disseminated intravascular
coagulation complicated by peripheral gangrene in a rhesus macaque (Macaca
mulatta) experimentally infected with Plasmodium coatneyi. Am
J Trop Med Hyg 76: 648-654.
- Moyano M, Mendez
F 2005. Defectos eritrociticos y densidad de la parasitemia en pacientes con
malaria por Plasmodium falciparum en Buenaventura, Colombia. Rev Panam
Salud Publica/Pan Am J Public Health 18: 25-32.
- Nakano Y, Fujioka
H, Luc KD, Rabbege JR, Todd GD, Collins WE, Aikawa M 1996. A correlation of
the sequestration rate of Plasmodium coatneyi-infected erythrocytes in
cerebral and subcutaneous tissues of a rhesus monkey. Am J Trop Med Hyg 55:
311-314.
- Nakazawa S, Looareesuwan
S, Fujioka H, Pongponratn E, Luc KD, Rabbege J, Aikawa M 1995. A correlation
between sequestered parasitized erythrocytes in subcutaneous tissue and cerebral
malaria. Am J Trop Med Hyg 53: 544-546.
- Navarro P, Baez
A, Vera Y, Figueira I, Garrido E, Martin A 2003. Paludismo como infeccion del
viajero adquirido en el Estado Sucre. Rev Fac Med (Caracas) 26: 34-38.
- Navarro P, Martin
A, Garrido E, Insausti C, Gutierrez H 1998. Palu-dismo en ninos: experiencia
del Hospital Universitario de Caracas. Arch Venez Pueric Pediatr 61:
109-112.
- Nguyen PH, Day
N, Pram TD, Ferguson DJ, White NJ 1995. Intraleucocytic malaria pigment and
prognosis in severe malaria. Trans R Soc Trop Med Hyg 89: 200-204.
- Noronha E, Costa
Alecrim M, Sierra Romero GA, Macêdo V 2000. Clinical study of falciparum malaria in children in Manaus, AM, Brazil. Rev Soc Bras Med Trop 33:
185-190.
- Obaldia N 2001. Evaluation of drug and vaccine candidates in the human malaria/Aotus monkey model, Panama, p. 1-94.
- Omar MS, Collins
WE, Contacos PG 1973. Gametocytocidal and sporontocidal effects of antimalarial
drugs on malaria parasites: I. Effect of single and multiple doses of primaquine
on Plasmodium cynomolgi. Exp Parasitol 34: 229-241.
- Osorio L, Gonzalez
I, Olliaro P, Taylor WRJ 2007. Artemisinin-based combination therapy for uncomplicated Plasmodium falciparum malaria in Colombia. Malaria J 6: 25.
- Othoro C, Lal AA,
Nahlen B, Koech D, Orago AS, Udhayakumar V 1999. A low interleukin-10 tumor
necrosis factor-alpha ratio is associated with malaria anemia in children residing
in a holoendemic malaria region in western Kenya. J Infect Dis 179: 279-282.
- Pérez Mato
S 1998. Anemia and malaria in a Yanomami Amerindian population from the southern
Venezuelan Amazon. Am J Trop Med Hyg 59: 998-1001.
- Perkins DJ, Weinberg
JB, Kremsner PG 2000. Reduced interleukin-12 and transforming growth factor-beta1
in severe childhood malaria: relationship of cytokine balance with disease severity. J Infect Dis 182: 988-992.
- Phillips RE, Looareesuwan
S, Warrell DA, Lee SH, Karbwang J, Warrell MJ, White NJ, Swasdichai C, Weatherall
DJ 1986. The importance of anaemia in cerebral and uncomplicated falciparum malaria: role of complications, dyserythropoiesis and iron sequestration. Q
J Med 58: 305-323.
- Price RN, Douglas
NM, Anstey NM 2009. New developments in Plasmodium vivax malaria: severe
disease and the rise of chloroquine resistance. Curr Opin Infect Dis 22:
430-435.
- Price RN, Simpson
JA, Nosten F, Luxemburger C, Hkirjaroen L, ter Kuile F, Chongsuphajaisiddhi
T, White NJ 2001. Factors contributing to anemia after uncomplicated falciparum malaria. Am J Trop Med Hyg 65: 614-622.
- Ramos Junior WM,
Sardinha JFJ, Costa MRF, Santana MS, Alecrim MGC, Lacerda MVG 2010. Clinical
aspects of hemolysis in patients with P. vivax malaria treated with primaquine
in the Brazilian Amazon. Braz J Infect Dis 14: 410-412.
- Rayner JC, Tran
TM, Corredor V, Huber CS, Barnwell JW, Galinski MR 2005. Dramatic difference
in diversity between Plasmodium falciparum and Plasmodium vivax reticulocyte binding-like genes. Am J Trop Med Hyg 72: 666-674.
- Restrepo-Pineda
E, Arango E, Maestre A, Do Rosario VE, Cravo P 2008. Studies on antimalarial
drug susceptibility in Colombia in relation to Pfmdr1 and Pfcrt. Parasitology
135: 547-553.
- Ritter K, Kuhlencord
A, Thomssen R, Bommer W 1993. Prolonged haemolytic anaemia in malaria and autoantibodies
against triosephosphate isomerase. Lancet 342: 1333-1334.
- Rodríguez-Morales
AJ, Ferrer MV, Barrera MA, Pacheco M, Daza V, Franco-Paredes C 2009. Imported
cases of malaria admitted to two hospitals of Margarita Island, Venezuela, 1998-2005. Travel Med Infect Dis 7: 44-48.
- Rodríguez-Morales
AJ, Sánchez E, Arria M, Vargas M, Piccolo C, Colina R, Franco-Paredes
C 2007. Haemoglobin and haematocrit: the threefold conversion is also non valid
for assessing anaemia in Plasmodium vivax malaria-endemic settings. Malaria
J 6: 166.
- Rodríguez-Morales
AJ, Sánchez E, Vargas M, Piccolo C, Colina R, Arria M 2006a. Anemia and
thrombocytopenia in children with Plasmodium vivax malaria. J Trop
Pediatr 52: 49-51.
- Rodríguez-Morales
AJ, Sanchez E, Vargas M, Piccolo C, Colina R, Arria M, Franco-Paredes C 2006b.
Pregnancy outcomes associated with Plasmodium vivax malaria in northeastern
Venezuela. Am J Trop Med Hyg 74: 755-757.
- Rodríguez-Morales
AJ, Sánchez E, Vargas M, Piccolo C, Colina R, Arria M, Franco-Paredes
C 2006c. Is anemia in Plasmodium vivax malaria more frequent and severe
than in Plasmodium falciparum? Am J Med 119: e9-10.
- Rodríguez-Morales
M, Brito Llano L, Fortun Prieto A, Diaz Dominguez MDLA, Montano Diaz MA, Rodriguez
Martinez S 1986. Anemia hemolitica por antipaludicos: estudio de 38 casos. Rev
Cuba Med 25: 609-613.
- Ronnefeldt F 1956.
Endemische malaria und anamie. Z Tropenmed Parasit 7: 48-63.
- Roshanravan B,
Kari E, Gilman RH, Cabrera L, Lee E, Metcalfe J, Calderon M, Lescano AG, Montenegro
SH, Calampa C, Vinetz JM 2003. Endemic malaria in the Peruvian Amazon Region
of Iquitos. Am J Trop Med Hyg 69: 45-52.
- Ruiz-Gil W, Tokeshi
Gusukuda-Shirota A, Pichilingue Prieto O 1994. Malaria por Plasmodium vivax:
aspectos clinicos y hematologicos. Rev Med Hered 5: 118-128.
- Sanchez Perovani
JA, Gonzalez Gonzalez BA, Valdes Dopaso LM 1983. Anemia hemolitica primaquinosensible. Rev Cuba Med 22: 1-4.
- Santana Filho FS,
Arcanjo AR, Chehuan YM, Costa MR, Martinez-Espinosa FE, Vieira JL, Barbosa MG,
Alecrim WD, Alecrim MG 2007. Chloroquine-resistant Plasmodium vivax,
Brazilian Amazon. Emerg Infect Dis 13: 1125-1126.
- Sarabia E, Ching
S, Unamuno M, Ramos A 1996. Paludismo y embarazo. Medicina (Guayaquil) 2:
129-132.
- Schmidt LH 1986.
Compatibility of relapse patterns of Plasmodium cynomolgi infections
in Rhesus monkeys with continuous cyclical development and hypnozoite
concepts of relapse. Am J Trop Med Hyg 35: 1077-1099.
- Sein KK, Brown
AE, Maeno Y, Smith CD, Corcoran KD, Hansukjariya P, Webster HK, Aikawa M 1993.
Sequestration pattern of parasitized erythrocytes in cerebrum, mid-brain, and
cerebellum of Plasmodium coatneyi-infected rhesus monkeys (Macaca
mulatta). Am J Trop Med Hyg 49: 513-519.
- Sempertegui F,
Estrella B, Toapanta FR, Torres DS, Calahorrano DE, Yeboah-Antwi K, Addo-Yobo
E, Arthur P, Newton S, Premji Z, Hubert M, Makwaya CS, Ssengooba F, Konde-Lule
J, Mukisa E, Hamer DH, MacLeod W, Duggan C, Fawzi W, Simon J, Mwanakasale V,
Mulenga M, Sukwa T, Tshiula J 2002. Effect of zinc on the treatment of Plasmodium
falciparum malaria in children: a randomized controlled trial. Am J Clin
Nutr 76: 805-812.
- Serghides L, Smith
TG, Patel SN, Kain KC 2003. CD36 and malaria: friends or foes? Trends Parasitol
19: 461-469.
- Shaffer N, Grau
GE, Hedberg K, Davachi F, Lyamba B, Hightower AW, Breman JG, Phuc ND 1991. Tumor
necrosis factor and severe malaria. J Infect Dis 163: 96-101.
- Silverman PH, Schooley
JC, Mahlmann LJ 1987. Murine malaria decreases hemopoietic stem cells. Blood
69: 408-413.
- Simpson JA, Silamut
K, Chotivanich K, Pukrittayakamee S, White NJ 1999. Red cell selectivity in
malaria: a study of multiple-infected erythrocytes. Trans R Soc Trop Med
Hyg 93: 165-168.
- Smith CD, Brown
AE, Nakazawa S, Fujioka H, Aikawa M 1996. Multi-organ erythrocyte sequestration
and ligand expression in rhesus monkeys infected with Plasmodium coatneyi malaria. Am J Trop Med Hyg 55: 379-383.
- Srichaikul T, Siriasawakul
T, Poshyachinda M 1976. Ferrokinetics in patients with malaria: haemoglobin
synthesis and normoblasts in vitro. Trans R Soc Trop Med Hyg 70:
244-246.
- Srichaikul T, Siriasawakul
T, Poshyachinda M, Poshyachinda V 1973. Ferrokinetics in patients with malaria:
normoblasts and iron incorporation in vitro. Am J Clin Path 60:
166-174.
- Stevenson MM, Su
Z, Sam H, Mohan K 2001. Modulation of host responses to blood-stage malaria
by interleukin-12: from therapy to adjuvant activity. Microbes Infect 3:
49-59.
- Stoute JA, Odindo
AO, Owuor BO, Mibei EK, Opollo MO, Waitumbi JN 2003. Loss of red blood cell-complement
regulatory proteins and increased levels of circulating immune complexes are
associated with severe malarial anemia. J Infect Dis 187: 522-525.
- Taverne J, Sheikh
N, de Souza JB, Playfair JH, Probert L, Kollias G 1994. Anaemia and resistance
to malaria in transgenic mice expressing human tumour necrosis factor. Immunology
82: 397-403.
- Tjitra E, Anstey
NM, Sugiarto P, Warikar N, Kenangalem E, Karyana M, Lampah DA, Price RN 2008.
Multidrug-resistant Plasmodium vivax associated with severe and fatal
malaria: a prospective study in Papua, Indonesia. PLoS Med 5: e128.
- Torres R JR, Magris
M, Villegas L, Torres V MA, Dominguez G 2000. Spur cell anaemia and acute haemolysis
in patients with hyperreactive malarious splenomegaly. Experience in an isolated
Yanomamo population of Venezuela. Acta Tropica 77: 257-262.
- Uscategui RM, Correa
AM, Carmona-Fonseca J 2009. Cambios en las concentraciones de retinol, hemoglobina
y ferritina en ninos paludicos colombianos. Biomedica 29: 270-281.
- Ventura AMRS, Pinto
AYN, Silva RSU, Calvosa VSP, Silva Filho MG, Souza JM 1999. Malaria por Plasmodium
vivax em crianças e adolescentes - aspectos epidemiologicos, clínicos
e laboratoriais. J Pediatr (Rio J) 75: 187-194.
- Vitor-Silva S,
Reyes-Lecca RC, Pinheiro TR, Lacerda MV 2009. Malaria is associated with poor
school performance in an endemic area of the Brazilian Amazon. Malar J 8:
230.
- Waitumbi JN, Opollo
MO, Muga RO, Misore AO, Stoute JA 2000. Red cell surface changes and erythrophagocytosis
in children with severe Plasmodium falciparum anemia. Blood 95:
1481-1486.
- Weatherall DJ,
Abdalla S, Pippard MJ 1983. The anaemia of Plasmodium falciparum malaria. Ciba Found Symp 94: 74-97.
- Weatherall DJ,
Miller LH, Baruch DI, Marsh K, Doumbo OK, Casals-Pascual C, Roberts DJ 2002.
Malaria and the red cell. Hematology Am Soc Hematol Educ Program: 35-57.
- Westenberger SJ,
McClean CM, Chattopadhyay R, Dharia NV, Carlton JM, Barnwell JW, Collins WE,
Hoffman SL, Zhou Y, Vinetz JM, Winzeler EA 2010. A systems-based analysis of Plasmodium vivax lifecycle transcription from human to mosquito. PLoS
Negl Trop Dis 4: e653.
- WHO - World Health
Organization 2000. Severe falciparum malaria. World Health Organization,
Communicable Diseases Cluster. Trans R Soc Trop Med Hyg 94 (Suppl. 1):
S1-90.
- WHO - World Health
Organization 2009. World Malaria Report 2009. Available from:
who.int/malaria/world_malaria_report_2009/en/index.html.
- WHO - World Health
Organization 2010. World Malaria Report 2010. Available from: who.int/malaria/world_malaria_report_2010/en/index.html.
- Wickramasinghe
SN, Abdalla SH 2000. Blood and bone marrow changes in malaria. Baillieres
Best Pract Res Clin Haematol 13: 277-299.
- Wickramasinghe
SN, Looareesuwan S, Nagachinta B, White NJ 1989. Dyserythropoiesis and ineffective
erythropoiesis in Plasmodium vivax malaria. Br J Haematol 72:
91-99.
- Yap GS, Stevenson
MM 1992. Plasmodium chabaudi AS: erythropoietic responses during infection
in resistant and susceptible mice. Exp Parasitol 75: 340-352.
- Zamora F, Ramírez
O, Vergara J, Arévalo-Herrera M, Herrera S 2005. Hemoglobin levels related
to days of illness, race and Plasmodium species in Colombian patients
with uncomplicated malaria. Am J Trop Med Hyg 73: 50-54.
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc11145t1.jpg]
[oc11145t3.jpg]
[oc11145t2.jpg]
|

{kind=link}
{kind=link}
{kind=link}