 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 130-133 Original Article Change in mutation patterns of Plasmodium vivax dihydrofolate reductase (Pvdhfr) and dihydropteroate synthase (Pvdhps) in P. vivax isolates from malaria endemic areas of Thailand Jiraporn KuesapI; Kanchana RungsrihirunratII; Pimwan ThongdeeI; Ronnatrai RuangweerayutIII; Kesara Na-BangchangI, + IPharmacology
and Toxicology Unit, Graduate Program in Biomedical Sciences, Thammasat University,
Pathumthani, Thailand Received 9 November
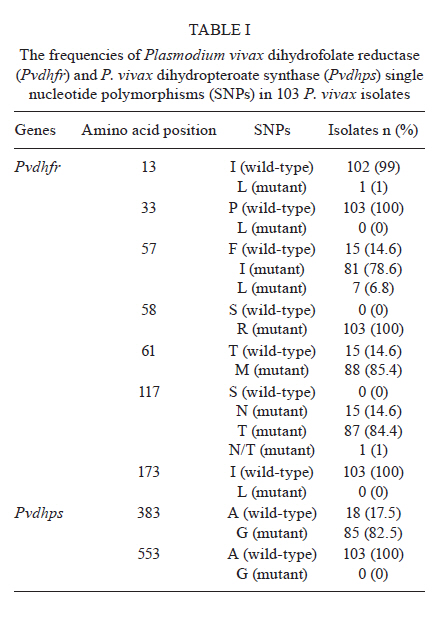
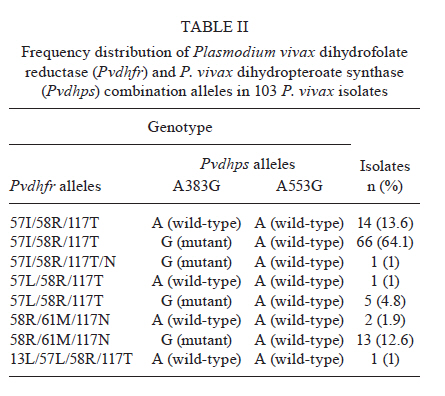
2010 Code Number: oc11150 Abstract Malaria is the most important public health problem in several countries. In Thailand, co-infections of Plasmodium vivax and Plasmodium falciparum are common. We examined the prevalence and patterns of mutations in P. vivax dihydrofolate reductase (Pvdhfr) and P. vivax dihydropteroate synthase (Pvdhps) in 103 blood samples collected from patients with P. vivax infection who had attended the malaria clinic in Mae Sot, Tak Province during 2009 and 2010. Using nested polymerase chain reaction-restriction fragment length polymorfism, we examined single nucleotide polymorphisms-haplotypes at amino acid positions 13, 33, 57, 58, 61, 117 and 173 of Pvdhfr and 383 and 553 of Pvdhps. All parasite isolates carried mutant Pvdhfr alleles, of which the most common alleles were triple mutants (99%). Eight different types of Pvdhfr and combination alleles were found, as follows: 57I/58R/117T, 57I/58R/117T, 57I/58R/117T/N, 57L/58R/117T, 57L/58R/117T, 58R/61M/117N, 58R/61M/117N and 13L/57L/58R/117T. The most common Pvdhfr alleles were 57I/58R/117T (77.7%), 57I/58R/117T/N (1%), 57L/58R/117T (5.8%) and 58R/61M/117N (14.5%). The most common Pvdhfr alleles were 57I/58R/117T (77.7%), 57I/58R/117T/N (1%), 57L/58R/117T (5.8%) and 58R/61M/117N (14.5%). Additionally, we recovered one isolate of a carrying a quadruple mutant allele, 13L/57L/58R/117T. The most prevalent Pvdhps allele was a single mutation in amino acid 383 (82.5%), followed by the wild-type A383/A553 (17.5%) allele. Results suggest that all P. vivax isolates in Thailand carry some combination of mutations in Pvdhfr and Pvdhps. Our findings demonstrate that development of new antifolate drugs effective against sulfadoxine-pyrimethamine-resistant P. vivax is required. Key words: Plasmodium vivax - Plasmodium vivax dihydrofolate reductase - Plasmodium vivax dihydropteroate synthase Introduction Antimalarial drug resistance is a major public health problem in tropical and sub-tropical countries including Thailand (MPH 2010). In Thailand, malaria is endemic throughout the country, with the highest incidence reported from Tak Province, the area along the Thai-Myanmar border (Na-Bangchang & Congpuong 2007). The antimalarial drug combination sulfadoxine-pyrimethamine (SP) (FansidarTM) was introduced into Thailand in 1972 as the first-line treatment for chloroquine resistant Plasmodium falciparum malaria. Soon after introduction of SP, resistance was reported along the Thai-Cambodian border, and by 1982, resistance was widespread throughout the country (Pinichpongse et al. 1982, Thaithong et al. 1992). Sulfadoxine and pyrimethamine target dihydropteroate synthase (dhps) and dihydrofolate reductase (dhfr) respectively; point mutations within the coding sequences of these genes are the primary cause of resistance to both drugs (Gregson & Plowe 2005). Although SP has never been used in Thailand to treat Plasmodium vivax infections, P. vivax is often co-infected with P. falciparum in relatively equal frequencies (Snounou & White 2004). Therefore, P. vivax has been unintentionally exposed to SP during treatment of P. falciparum infection, resulting in progressive selection of SP-resistant P. vivax (Imwong et al. 2003). Molecular studies revealed that point mutations in the dhfr and dhps genes of both P. falciparum and P. vivax parasites confer resistance to SP (Imwong et al. 2001, Gregson & Plowe 2005). The distribution of mutant alleles of these genes varies among different geographical regions and is related to the intensity of SP use for treatment of P. falciparum. The prevalence of P. vivax dhfr (Pvdhfr) and dhps (Pvdhps) mutations have been investigated in several malaria endemic regions including Indonesia, India, Sri Lanka, Afghanistan, Iran, Papua New Guinea, Madagascar and Thailand. Over 20 different Pvdhfr alleles have now been described (Hawkins et al. 2007). Studies in Thailand and India showed that mutations at Pvdhfr amino acids 57, 58, 61, 117 and 173 (Imwong et al. 2003, Barnadas et al. 2008) were found to be associated with clinical antifolate resistance (Imwong et al. 2001, Hawkins et al. 2007). In Pvdhps, five mutations have been identified at amino acids 382, 383, 512, 553 and 585 (Hawkins et al. 2007, Barnadas et al. 2008, Rungsihirunrat et al. 2008). In this paper, we determine the prevalence and diversity of Pvdhfr and Pvdhps mutant alleles in P. vivax isolates collected from Mae Sot District, an endemic area of Thailand along the Thai-Myanmar border. This information will assist in the development of effective new antifolates. Subjects, Materials and Methods Study areas and sample collection - A total of 103 blood samples with P. vivax mono-infection were collected from patients attending malaria clinics in Mae Sot during 2009 and 2010. Approval for the study protocol was obtained from the Ethics Committees of Mae Sot General Hospital, Tak Province. Blood samples of between 200-300 µL were collected by finger-prick onto filter paper (Whatman 3). Dried filter paper samples were stored in small zipper plastic bags prior to the extraction of parasite DNA for analysis by polymerase chain reaction (PCR). Giemsa-stained thin and thick blood smears were prepared and examined microscopically for the presence of P. vivax parasites. Extraction of parasite genomic DNA - Parasite genomic DNA was extracted from individual dried blood spots on filter paper using the QIAamp DNA Extraction Mini-kit (QIAGEN) and used as the template for PCR amplification. Amplification of Pvdhfr - Point mutations of Pvdhfr in all P. vivax isolates were investigated by nested PCR-restriction fragment length polymorfism (PCR-RFLP) at seven amino acid positions (13, 33, 57, 58, 61, 117 and 173) according to previously described methods (Imwong et al. 2001, 2003). Amplification of Pvdhps - Point mutations of Pvdhps at two amino acid (383 and 553) in all P. vivax isolates were investigated by nested PCR-RFLP (Rungsihirunrat et al. 2008). Results Detection of mutations in the Pvdhfr and Pvdhps genes - The frequencies of Pvdhfr and Pvdhps mutations are summarised in Table I. Point mutations in Pvdhfr were detected in five of seven amino acids investigated (13, 57, 58, 61 and 117), whereas wild-type alleles were detected at two amino acid positions (33 and 173). All isolates carried mutations at amino acids 58 (58R) and 117 (117N, 117T, 117N/T). One (1%) and 88 (85.4%) isolates carried mutations at amino acids 13 (13L) and 61 (61M), respectively. We recovered two mutations at amino acid 57, 57I and 57L. For Pvdhps, the most prevalent allele recovered was 383G (82.5%) and the wild-type A383/A553 (17.5%) alleles. Distribution of Pvdhfr and Pvdhps combination alleles - Table II summarises the distribution of eight different combinations of alleles recovered from our isolates of Pvdhfr (5 different alleles) and Pvdhps (2 different alleles). The most common Pvdhfr alleles were the triple mutants 57I/58R/117T (77.7%), 57I/58R/117T/N (1%), 57L/58R/117T (5.8%) and 58R/61M/117N (14.5%); only one isolate carried a quadruple mutation (13L/57L/58R/117T). The most prevalent genotype recovered was the triple mutant Pvdhfr 57I/58R/117T with the single mutant Pvdhps 383G (64.1%), followed by the triple mutant Pvdhfr 57I/58R/117T with a wild-type Pvdhps allele (13.6%) and finally the triple mutant Pvdhfr 58R/61M/117N with the single mutant Pvdhps 383G (12.6%). We did not recover any isolates that carried wild-type alleles of both genes. Discussion In 2005, we conducted a study in the same area (Mae Sot District) and collected a total of 32 isolates that were found to contain a combination of Pvdhfr mutant alleles (81.3% quadruple, 12.5% triple, 6.2% double) with a single or double mutant allele of Pvdhps (codons 383 and 553) (Rungsihirunrat et al. 2007). In the earlier study and in the study presented here, we did not detect any isolate carrying wild-type alleles of both genes. In fact, mutations at amino acids 58 and 117 of Pvdhfr were found in 100% of isolates. Another recent study of P. vivax isolates from the same area found that 57.1% of all isolates contained a 57I/58R/61M/117T triple mutation in Pvdhfr and a 383G/553G double mutation in Pvdhps (Lu et al. 2010). Of the five Pvdhfr amino acids (57, 58, 61, 117, 173) under investigation in our study, single (57L), double (58R, 117N) and quadruple (57I/L, 58R, 61M, 117T) mutations were found in 3.6%, 10.7% and 85.7% of the isolates, respectively. In Pvdhps sequences, wild-type, single (383G) and double (383G and 553G) mutant alleles were found at frequencies of 3.6%, 14.3% and 82.1%, respectively. All these data suggest that prevalence and patterns of mutant Pvdhfr and Pvdhps alleles in P. vivax samples isolated from the same area may vary over time in response to changing degrees of drug pressure. In Mae Sot, P. vivax and P. falciparum co-infect in an equal ratio. SP was used as the first-line treatment for P. falciparum in Thailand from 1972-1982 leading to a progressive selection of SP-resistant alleles in P.vivax (Imwong et al. 2003). Monitoring of P. vivax isolate drug sensitivity in vitro (Russell et al. 2003, Kosaisavee et al. 2006) in parallel with genotyping for markers of antimalarial resistance is essential for guiding effective treatment of P. vivax infection with antifolate drugs. In vitro sensitivity data obtained from a yeast expression system has been shown to provide correlated sensitivity data for P. vivax and may be more applicable for this purpose (Rungsihirunrat et al. 2007). We observed an increase in geometric mean IC50 values of antifolates with an increasing number of Pvdhfr mutations, from double to quadruple (Rungsihirunrat et al. 2007). Quadruple mutant alleles confer decreased sensitivity to pyrimethamine but retain sensitivity to WR99210 (Rungsihirunrat et al. 2007). Further examination of the association between different patterns of Pvdhfr and Pvdhps mutation and in vitro sensitivity of P. vivax isolates using the yeast expression system is needed. Several studies have investigated the prevalence and diversity of Pvdhfr and Pvdhps in P. vivax isolates from different malaria endemic areas of Thailand. In two studies conducted during 1992-1998 (Imwong et al. 2001, 2005) of patients admitted to the Bangkok Hospital for Tropical Diseases, double and triple Pvdhfr mutant alleles (amino acids 57, 58, 117) were found in almost all (n = 99,99%) of the isolates collected, the majority of which consisted of the triple mutant 57L/58R/117N allele (53%) (Imwong et al. 2001). Wild-type, single and double mutant alleles of amino acids 383 and 553 in Pvdhps were detected among 33 isolates (Imwong et al. 2005). Nine isolates carried four (57L/58R/61M/117T) or more (13L/57L/58R/61M/117T) point mutations in Pvdhfr, together with two point mutations in Pvdhps (Imwong et al. 2005). Previously, we described a collection of 160 isolates from 10 malaria endemic areas (Thai-Myanmar border, 117 isolates, Thai-Cambodian border, 13 isolates, and Thai-Malaysian border, 30 isolates) in which we detected wild-type, single, double, triple and quadruple mutant Pvdhfr alleles in 1.9%, 0.6%, 35.6%, 2.5% and 59.4% of all isolates, respectively. Only 1.3% of isolates were wild-type Pvdhps at amino acids 383 and 553 while single mutants at amino acid 383 corresponded to 26.9% and double mutant alleles at both amino acids corresponded to 60% of all isolates. The prevalence and patterns of Pvdhfr and Pvdhps mutations vary not only over time in the same malaria endemic area, but also across geographical areas. The common Pvdhfr alleles 58R and 117T/N were found in the southeast Asian countries of East Timor (de Almeida et al. 2010), Thailand (Imwong et al. 2001, Lu et al. 2010), Myanmar (Lu et al. 2010), Vietnam and the Philippines (Auliff et al. 2006), in India (Kaur et al. 2006, Alam et al. 2007), in the African countries of French Guinea (Barnadas et al. 2009) and Madagascar (Barnadas et al. 2008). Mutants in Pvdhps at amino acids 383 and 553 were found at low prevalence in most geographic regions including East Timor (de Almeida et al. 2010), Korea (Lu et al. 2010), Iran (Zakeri et al. 2009) and Pakistan (Zakeri et al. 2011), whereas they were found at high prevalence in Thailand (Imwong et al. 2005, Rungsihirunrat et al. 2008). Isolates carrying Pvdhps double mutations co-existing with multiple mutations (4 or more amino acids) of Pvdhfr are being reported with increasing frequency (Imwong et al. 2005). Moreover, quadruple mutant alleles of Pvdhfr associated with SP treatment failure were reported in Myanmar (Na et al. 2005), India (Kaur et al. 2006), Papua New Guinea, Vanuatu (Auliff et al. 2006) and Thailand (Rungsihirunrat et al. 2007). Increasing treatment failure with SP for uncomplicated P. falciparum malaria in several areas has led to the development of new combinations of antifolates, sulfas and dihydrofolate inhibitors such as LapDapTM and WR99210. In vitro sensitivity data has revealed that the quadruple mutant alleles of Pvdhfr confer resistance of P. vivax to pyrimethamine but retain sensitivity to WR99210 (Rungsihirunrat et al. 2007). References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11150t1.jpg] [oc11150t2.jpg] |
| |||||||||

{kind=link}
{kind=link}