 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
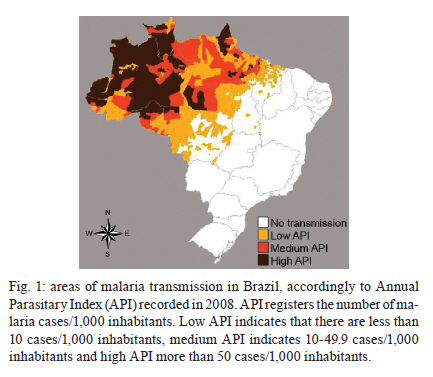
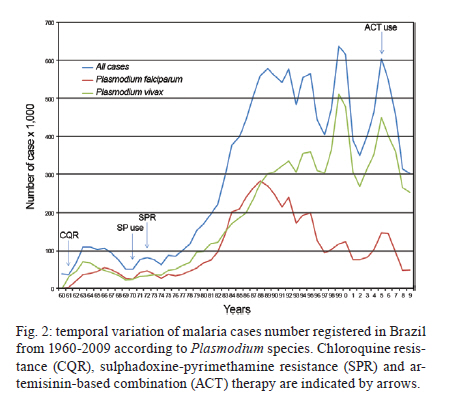
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 159-166 Original Article Chemoresistance of Plasmodium falciparum and Plasmodium vivax parasites in Brazil: consequences on disease morbidity and control Bianca Ervatti GamaI; Marcus Vinicius Guimarães LacerdaII; Cláudio Tadeu Daniel-RibeiroI; Maria de Fátima Ferreira-da-CruzI, + ILaboratório
de Pesquisas em Malária, Centro de Pesquisa, Diagnóstico e Treinamento
em Malária, Ministério da Saúde, Instituto Oswaldo Cruz-Fiocruz,
Av. Brasil 4365, 21040-360 Rio de Janeiro, RJ, Brasil Key words: Plasmodium falciparum - Plasmodium vivax - malaria - chemoresistance - Brazil Malaria, one of the most challenging infectious diseases, is an important global public health problem. This disease is endemic in 106 countries and there are reports of about 225 million cases and 781,000 deaths worldwide per year (WHO 2010). In Brazil, despite intensive control efforts, malaria remains clinically important cause of febrile syndrome in local populations and in travellers in areas with endemic transmission, particularly in the Amazon Basin (Oliveira-Ferreira et al. 2010). The Brazilian Amazon comprises nine states: Amazonas (AM), Acre (AC), Roraima (RR), Amapá (AP), Pará (PA), Rondônia (RO), Tocantins (TO), Mato Grosso (MT), and part of Maranhão (MA).In the 1940s, it was estimated that six million people were infected with malaria per year across Brazil, corresponding to roughly 20% of the national population. A nationwide campaign for malaria eradication was launched in 1965, as recommended by the World Health Organization (WHO). This campaign, based on indoor dichlorodiphenyltrichloroethane (from its trivial name, DDT) spraying and chloroquine (CQ) treatment for febrile cases, curtailed malaria transmission in the majority of the country (Loiola et al. 2002), and transmission areas were subsequently limited to the north of the country (Fig. 1). The first report of resistance to an antimalarial drug in Brazil was in 1907. This report was presented anecdotally at a Congress and described the inefficacy of quinine (QN) prophylaxis in workers during the construction of the Madeira-Mamoré Railway in RO. In 1908, Miguel Couto also reported falciparum malaria patients not responding to QN in Rio de Janeiro (RJ), but it was Arthur Neiva (Neiva 1910) who published the first scientific evidence of QN resistance in workers from RJ two years later. Five decades later, two initial foci of CQ-resistant Plasmodium falciparum parasites were reported in Colombia (Moore & Lanier 1961, Young & Moore 1961) and on the frontier between Thailand and Cambodia (Harinasuta et al. 1965). The resistant strains gradually spread throughout Southeast Asia and South America. In Brazil, the first report of CQ-resistant P. falciparum cases was in 1961. The famous physician, José Rodrigues da Silva, from Federal University of Rio de Janeiro, identified resistance in individuals returning from road construction in the states of Bahia, RO, PA and MA (Silva 1961), but these findings did not have any repercussion. Starting in the mid-1960s, the Brazilian government launched and sponsored a massive colonisation project of the northern region in the Amazon Basin. This rapid and uncontrolled migratory movement of non-immune subjects (Daniel-Ribeiro et al. 2008), together with the emergence and expansion of CQ-resistant strains of P. falciparum to other Brazilian states such as AC, RR, AM (Ferraroni et al. 1981), MT and AP (Reyes 1981), led to a 10-fold increase in the incidence of malaria-induced morbidity between 1970 and the mid-1980s. Fortunately, malaria was almost entirely absent from the non-Amazon areas of Brazil at this time. Expansion of P. falciparum CQ chemoresistance, the introduction of sulphadoxine-pyrimethamine (SP) combination and the spread of resistance - Following the appearance of CQ resistance (CQR) in Brazil in the 1960s, other countries in South America and Asia (Clyde 1960, Comer et al. 1968) began reporting the occurrence of CQ-resistant P. falciparum parasites and simultaneously, amodiaquine (AQ)-resistant parasites. This reinforced the call to action for all of the control programs worldwide, as they were now facing the ghost of drug resistance (Souza 1992). The spread of CQ-resistant strains (Box et al. 1963, Silva & Lopes 1964) accelerated the search for new drugs and SP was highlighted as the new option for falciparum malaria treatment in the 1970s after a clinical trial that showed a high cure rate of CQ-resistant parasites treated with this drug combination (Walker & Lopez-Antunano 1968). Fatefully, the appearance of SP-resistant P. falciparum parasites was verified in Brazilian isolates from TO (Almeida Netto et al. 1972) within a short period of time. Soon after, a high degree of SP-chemoresistance in other states from Brazil was reported (Alecrim et al. 1982a, b, Souza 1983), leading to a climax of at least 90% SP resistance (SPR) in 1987 (Souza 1992). By the end of the 1980s, treatment failure with CQ was increasing, reaching 100% in some situations (Boulos et al. 1986, Kremsner et al. 1989, Andrade et al. 1992, Couto et al. 1995, Segurado et al. 1997, Póvoa et al. 1998, Zalis et al. 1998, Cerutti et al. 1999b). Policy treatment changes - By the 1980s, CQ-resistant and SP-resistant P. falciparum strains were contributing to the remarkable increase in morbidity (Fig. 2) and, in response to this drug resistance, it was urged that QN should be reintroduced as standard therapy in 1984 in association with tetracycline (Boulos et al. 1986). In 1986, a new initiative for malaria control and morbidity reduction led to the introduction of other drugs for malarial therapy. This included AQ (Kremsner et al. 1987) and mefloquine (MQ) (Loiola et al. 2002) for the treatment of drug-resistant P. falciparum. However, a reduction of the in vivo and in vitro MQ susceptibility (Alecrim 1981) and an in vitro MQ resistance level of 27.5% had been reported before the official introduction of the drug in 1987 (Boulos et al. 1986). In contrast, some clinical trials (Souza 1983, 1986, Souza et al. 1985) revealed a 100% cure rate and MQ resistance was not detected by subsequent in vitro studies (Póvoa et al. 1998, Zalis et al. 1998). This contrasted with the previously detected reduction of susceptibility to MQ in vitro (Couto et al. 1995). In 1999, however, Cerutti et al. (1999b) once again recognised the reduction of susceptibility to MQ, in addition to observing a case of in vivo resistance (Cerutti et al. 1999a). In the next two years, high levels of MQ resistance were seen in children from AM (Noronha et al. 2000) and in patients from PA (Calvosa et al. 2001). Concerning AQ, in vivo resistance was also detected at multiple Brazilian localities very soon after its introduction (Reyes et al. 1986, Passos et al. 1987) and high-grade in vitro resistance was rapidly verified (Kremsner et al. 1989). In 2001, the Brazilian National Malaria Control Program instituted the combination of QN plus doxycycline, followed by primaquine (PQ) as a gametocytocidal drug (0.75 mg/kg in a single dose) as a first-line drug combination, and MQ plus PQ was introduced as a second-line drug combination. It is important to remark that QN was reintroduced as a first-line drug, despite studies in the 1980s that showed the parasites' in vitro chemoresistance to this drug (Souza et al. 1985, Boulos et al. 1986, Souza 1986, Kremsner et al. 1989, Cerutti et al. 1999b, Calvosa et al. 2001). This occurred because WHO had recently recommended the use of drug combinations, instead of single drug therapy, due to the fast ability of P. falciparum parasites to develop resistance to individual drugs. QN usage had been instituted in association with tetracycline, a class of antibiotics with slow schizonticidal action, but with a longer half-life than QN. Tetracycline/doxycycline were chosen over clindamycin for association with QN for the following two reasons. First, tetracycline/doxycycline have a lower cost than clindamycin and, second, a large batch of these drugs had been acquired for the treatment of patients with cholera during the outbreak in AM and were therefore available for use (MGC Alecrim, unpublished observations). As might have been predicted, a continuous increase in QN resistance resulted in a pronounced P. falciparum morbidity intensification, and a phase II study was conducted in the Brazilian Amazon to compare treatments with artemether/lumefantrine (Coartem®, supplied by Novartis) to QN/doxycycline (Alecrim et al. 2006). In view of the promising results, the artemisinin-combination therapy (ACT) (represented by Coartem) and artesunate plus MQ (supplied by Farmanguinhos) were introduced by the Brazilian Malaria Control Program for uncomplicated falciparum malaria treatment starting in 2007 and 2008, respectively (Ladislau 2008, Carmargo et al. 2009). Since the end of the 1990s, parenteral artesunate or artemether had been in use in association with clindamycin for severe malaria treatment. In adopting ACTs as the first line of treatment for non-severe falciparum malaria, Brazil abolished the systematic use of PQ as a gametocytocidal drug for P. falciparum. The only exception involved cases in which gametocytes were observed in the thick blood smear. This decision should be reviewed, because PQ use substantially decreases the carriage of gametocytes (Smithuis et al. 2010), a fact that certainly should have an impact on reducing malaria transmission, and it was demonstrated that PQ is safe to use in glucose-6-phosphate-dehydrogenase (G6PD)-deficient children using the preconised prescript dose (Shekalaghe et al. 2007). At this time, P. falciparum field isolates from French Guyana and Senegal with reduced in vitro sensitivity to artemether (Jambou et al. 2005), as well as parasites from Cambodia, which exhibit slightly reduced in vitro sensitivity with significant prolongation of parasite clearance times after artesunate therapy have been reported (Noedl et al. 2008, Dondorp et al. 2009). However, no artemisinin derivatives treatment failures have been reported in Brazil. Thus, resistance surveillance, with the support of in vitro studies in sentinel institutions, is essential to predict new policy interventions in the near future. Emergence of P. vivax chemoresistance - In Brazil, as in many parts of the world, Plasmodium vivax developed resistance to CQ much later than did P. falciparum. One possible explanation is that P. vivax gametocytes develop much earlier during the course of the disease, when no drug pressure is expected to occur. In the case of P. falciparum, gametocytes circulate in the peripheral blood and are accessible to mosquitoes only seven days after disease onset. This not only substantially facilitates parasite control but also results in the selection of gametocytes resistant to the drugs used for therapy. Studying CQ-resistant P. vivax is complicated, even in non-endemic areas due to the possibility of relapses originating from hypnozoites in the liver making it difficult to recognise chemoresistant parasites. In view of this, CQR is acknowledged when there is parasite recurrence in the peripheral blood during the first 28 days after the beginning of CQ therapy, at which time no parasites should be circulating because of the presence of therapeutic drug levels. Ideally, the plasma levels of CQ and its active metabolite, desethylchloroquine, should be dosed to withdraw therapeutic failure and therefore, to institute chemoresistance (Baird 2004, 2009). In 1992 two Italian authors claimed to have identified the first case of CQR in Brazil (Garavelli & Corti 1992). Soon after, however, it became clear that it was not a case of a true recrudescence (Loyola & Rodriguez 1992). The first reliable case of in vivo CQR in Brazil was from a patient treated at the Tropical Medicine Foundation Dr Heitor Vieira Dourado in Manaus, AM (Alecrim et al. 1999a). These P. vivax parasites were also resistant to MQ. Subsequently, the same group also reported a P. vivax strain resistant to PQ (a drug used along with CQ for the radical cure of liver stages) and CQ (Alecrim et al. 1999b). Further evidence of P. vivax CQR appeared in 2007, also in Manaus. A proper 28-day follow-up and plasmatic drug dosage was conducted in 109 patients and 10% of the infections were found to be drug-resistant (Santana Filho et al. 2007). Interestingly, this resistance did not seem to spread throughout the Amazon Region; in a similar study in AC, no CQR was detected (Orjuela-Sánchez et al. 2009a). It has been claimed that the concomitant use of PQ and CQ could result in a synergistic effect against asexual forms of P. vivax, as has been shown in P. falciparum parasites (Bray et al. 2005). However, the few in vivo data available for P. vivax show that the parasitological response to CQ and to CQ plus PQ is quite similar (Pukrittayakamee et al. 1994). Consequently, the possibility that such synergism results in the underreporting of CQR in areas where PQ is systematically used should be examined. It has been hypothesised that severe vivax malaria disease could be related to CQR (Tjitra et al. 2008). Data from Brazil suggest that this relationship would not be straightforward (Alexandre et al. 2010). However, the fact that CQR has increased in many parts of Southeast Asia suggests that CQ should no longer be used routinely. ACTs are the major alternative drugs used to treat P. vivax, despite the scarcity of evidence on their efficacy against P. vivax parasites (Douglas et al. 2010). In relation to PQ resistance, it is well-known that the WHO recommended a dose of 0.25 mg/kg/day for 14 days which differs from the abbreviated regimen of 0.5 mg/kg/day for seven days (Galappaththy et al. 2007). However, in the mid-1990s, the Brazilian Malaria Control Program changed its policy and recommended the abbreviated regimen due to better compliance and tolerability, despite a slightly inferior efficacy (Krudsood et al. 2008). In spite of the methodological limitation of studying relapses in endemic areas, hypnozoite resistance to PQ is being increasingly reported and in many countries higher doses of PQ are needed for radical cure. It is worth noting that most of these studies had been performed without supervised administration of the drug, which could generate an overestimation of the number of cases of relapses (Baird & Rieckmann 2003). In Brazil, one robust study was performed outside the transmission area with supervised treatment of malaria vivax with CQ plus PQ. This study showed a PQ failure rate as high as 24.5% as determined by relapses, which primarily occurred after three months (Boulos et al. 1991). Another issue that deserves consideration is PQ dosage in patients with G6PD-deficiency. This issue is important because resistance to PQ is dose-dependent and the A- human genotype, the most frequent in Brazil, is associated with severe haemolysis in a dose-dependent manner (Santana et al. 2009, Ramos Junior et al. 2010). In addition, screening for G6PD is not performed routinely before antimalarial treatment in Brazil. The efficaciously elimination of the hypnozoite is a major concern for malaria control because relapses may contribute enormously to the burden of the disease (Wells et al. 2010). Presently, only PQ is available for the radical cure and very few compounds are under pre-clinical investigation due to the difficulties of an experimental or in vitro model for large-scale drug screening. Tafenoquine is a promising drug with good preliminary data on relapse prevention and with a more convenient posological administration. Unfortunately, the study of vivax malaria has been relatively neglected until recently. Even in areas where it predominates, local scientific groups concentrate their research on P. falciparum parasites (Lacerda et al. 2007). P. falciparum and P. vivax surveillance studies - Since the Amsterdam Conference at the beginning of the 1990s, early and effective treatment of malaria has been considered the cornerstone of malaria control. Plasmodium resistance to antimalarials is one of the major obstacles to achieving this goal. A systematic, comprehensive and up-to-date monitoring of resistance is important to guide malaria treatment policies, to detect changing patterns early and to avoid the spread of resistance. Generally, three main approaches are used to study and/or measure antimalarial drug resistance for P. falciparum: (i) classical, gold-standard, in vivo follow-up based on clinical and parasitological outcomes obtained in an standard period of time, coupled to complementary analysis to determine the origin of recurrent parasitemia, (ii) an in vitro test based on culturing malaria parasites to measure their intrinsic sensitivity and trends in resistance to antimalarial drugs and, as a modern public health tool, (iii) molecular genotyping of markers to identify genetic mutations that were previously associated with antimalarial drug resistance. However, for P. vivax, the readily obtainable methodologies are elusive. Recognition of resistant parasites is challenging and no consistent, long-term, in vitro method for culturing P. vivax parasites is available. Additionally, no trustworthy molecular marker for CQR has been identified thus far. In Brazil, P. falciparum in vitro tests have been performed since 1981 either alone (Ferraroni et al. 1981, Reyes 1981, Couto et al. 1995, Calvosa et al. 2001, Menezes et al. 2001), in conjunction with in vivo follow-up studies (Boulos et al. 1986, Neifer & Kremsner 1991, Segurado et al. 1997, Cerutti et al. 1999b) or along with molecular genotyping (Póvoa et al. 1998, Zalis et al. 1998, Vieira et al. 2001, Ferreira et al. 2008). As far as P. falciparum molecular genotyping is concerned, the first informed assay identified only one mutation or single-nucleotide-polymorphism (SNP) in the pfdhfr gene that was associated with pyrimethamine resistance (Peterson et al. 1991). When complete genotyping was performed in Brazil (Vasconcelos et al. 2000), a high number of SNPs were identified in critical codons of the pfdhfr (pyrimethamine) and pfdhps (sulphadoxine) genes, and these were related to acquisition of full SPR (Cowman et al. 1988, Peterson et al. 1988, Zolg et al. 1989, Brooks et al. 1994). In the case of CQ molecular resistance, two studies published in 1998 presented SNPs of the pfmdr1 gene (Póvoa et al. 1998, Zalis et al. 1998), which were associated with a modified response to CQ and to other structurally unrelated antimalarials (Duraisingh & Cowman 2005). Years later, when resistance to CQ was also ascribed to the gene that encodes a transmembrane transport protein named pfcrt (Fidock et al. 2000), two studies that genotyped the major codons of this gene found that all the isolates presented CQ-resistant haplotypes (Vieira et al. 2001, 2004). This supported the usefulness of this gene as a molecular marker of in vivo CQR. More recently, a decrease in CQ-sensitive P. falciparum parasites, as well as the downturn of triple mutants associated with SPR, were identified in areas where drug pressure had been removed for a substantial period of time (Kublin et al. 2003, Mita et al. 2003, Wang et al. 2005, Zhou et al. 2008, Bacon et al. 2009). In Brazil, a study examining CQ and SP markers revealed, for the first time in the country, that P. falciparum sensitive strains can also circulate in parasite populations, at least in PA (Gama et al. 2009b). Soon after, in response to the potential development of artemisinin resistance, two studies were published. In the first investigation, the presence of mutations on artemisinin putative resistance-associated genes pfcrt, pfmdr1 and pfatpase6, the absence of a pfatpase6 chief mutation and satisfactory artemether and artesunate in vitro activities were reported (Ferreira et al. 2008). In the other study, known polymorphisms at pfmdr1 and pfatpase6 were identified in samples collected from three Brazilian states before the introduction of ACTs in Brazil. This assembled a genetic snapshot of P. falciparum parasites that could be used as a baseline reference for future studies (Gama et al. 2010). With respect to P. vivax molecular genotyping, orthologous P. falciparum genes potentially linked to CQ chemoresistance, such as pvcrt-o and pvmdr1, have been identified (Nomura et al. 2001, Brega et al. 2005). Molecular characterisation of P. vivax isolates was also carried out in Brazil (Sá et al. 2005, Gama et al. 2009a, Orjuela-Sánchez et al. 2009b). In these publications, the sequence of pvmdr1 in three Brazilian isolates was shown, without association between mutations and in vitro outcomes (Sá et al. 2005). Complete analysis of coding and noncoding sequences of the putative resistance genes pvcrt-o and pvmdr1 was conducted in seven samples and no association with CQR was found (Orjuela-Sánchez et al. 2009b). The last survey examined the relationship between SPR and the pvmdr1 and pvdhfr genes in a set of 28 samples (de Pécoulas et al. 1998, Eldin de Pécoulas et al. 1998). This identified a potential leader mutation in pvmdr1 in 100% of the examined isolates (Gama et al. 2009a). This study contrasted with the reported 10% of in vivo CQ treatment failures (Santana Filho et al. 2007). Moreover, the presence of mutations in pvdhfr gene could reflect the selection of SP-resistant P. vivax parasites due to misdiagnosed, or even undiagnosed, P. falciparum and P. vivax co-infections (Gama et al. 2009a). South-American surveillance network - Amazon Network for the Surveillance of Antimalarial Drug Resistance (RAVREDA) - In 2001, a surveillance network across the Amazon and South American countries was created in response to the challenge of antimalarial drug resistance in the Amazon. It was called RAVREDA and it has received financial support from the US Agency for International Development and the Pan-American Health Organization. RAVREDA activities formally began in 2002. Since then, several studies have been performed in Bolivia, Brazil, Colombia, Ecuador, Guyana, French Guiana, Peru, Suriname and Venezuela. The main objectives are to strengthen resistance surveillance and to furnish an evidence-based formulation of drug policies to promote the suitable use of antimalarials. Evaluation of the therapeutic effectiveness of antimalarials is performed utilising in vivo as well as in vitro efficacy studies. These studies apply a WHO standardised protocol to a sample of patients seeking malaria diagnostic and care services in their own health system. These areas of care services are then identified as sentinel sites. Through 2005, the network has performed 81 studies in South America and has generated information on therapeutic effectiveness that has guided changes in therapeutic regimens in several South American countries (PAHO 2007). Although the majority of RAVREDA's past activities have concerned P. falciparum, one must consider that, in 2004, P. vivax accounted for more than 70% of all cases of malaria in Latin America (PAHO 2006) and CQ-resistant strains have been reported in Guyana, Peru and Colombia (Soto et al. 2001, Ruebush et al. 2003, Esterre et al. 2009). In Brazil, besides the initial report by Alecrim et al. (1999a) and the report from Santana Filho et al. (2007), another unpublished RAVREDA study performed in the municipality of Careiro (AM), found that 18% of P. vivax infections were CQ-resistant. This was based on determination of the serum levels of both CQ and desethylchloroquine (PAHO 2006). Brazil is a continental country with approximately 8.5 million square miles and 47% of South America's landmass. It is responsible for about 56% of the total malaria cases in the Americas (PAHO 2008). In such a large territory with a huge burden of disease, antimalarial drug surveillance is crucial to maintain an efficient control program. The data presented herein show that drug resistance surveillance assays must be conducted on a regular basis to assess antimalarial efficacy and to ensure that the information is available to policy makers. At the present time, P. falciparum surveillance studies should investigate artemisinin, as well as its counterparts in ACT formulation; for P. vivax, the actual amount of CQR needs to be determined. Additionally, RAVREDA efforts should be strengthened to allow for easy disclosure of the obtained results. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11153f2.jpg] [oc11153f1.jpg] |
| |||||||||

{kind=link}
{kind=link}