 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 193-201 Original Article Synergism/complementarity of recombinant adenoviral vectors and other vaccination platforms during induction of protective immunity against malaria Ana Paula Morais Martins Almeida; Oscar Bruna-Romero+ Departamento de
Microbiologia, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brasil Received 1 April
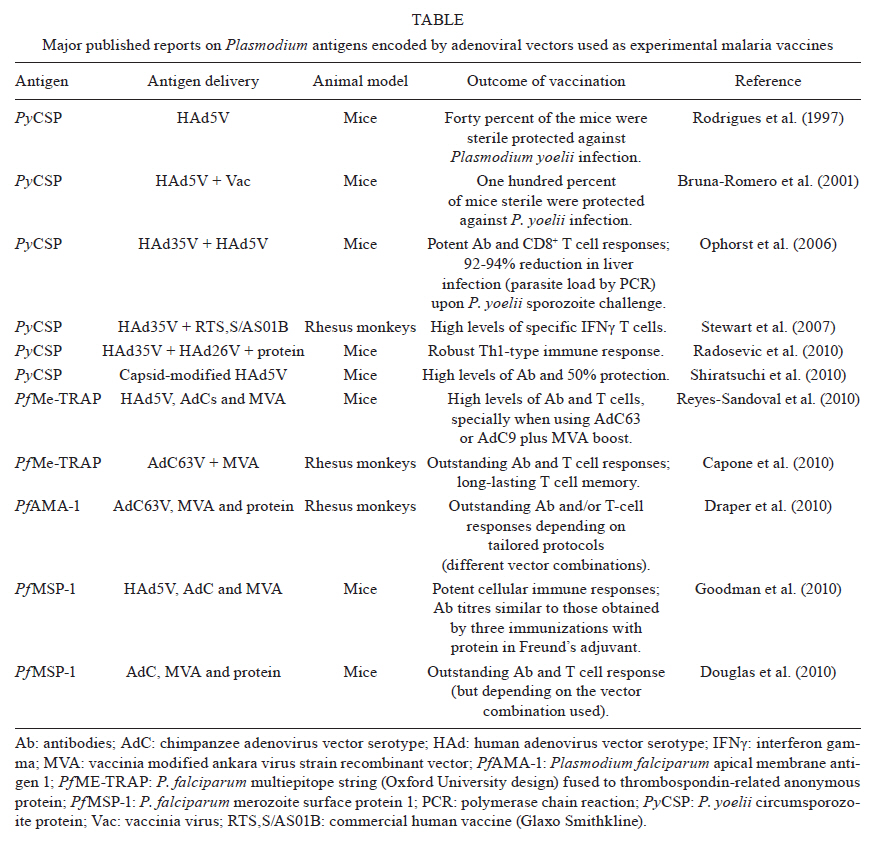
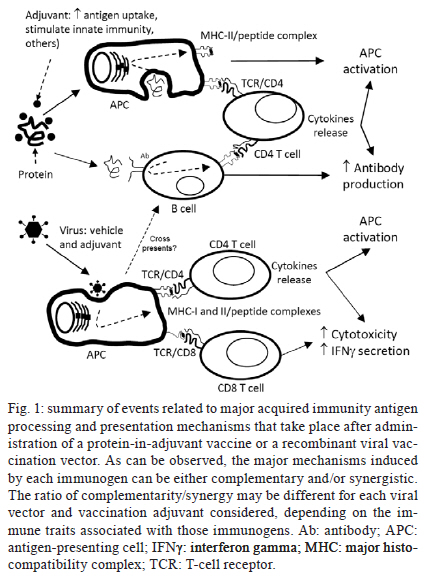
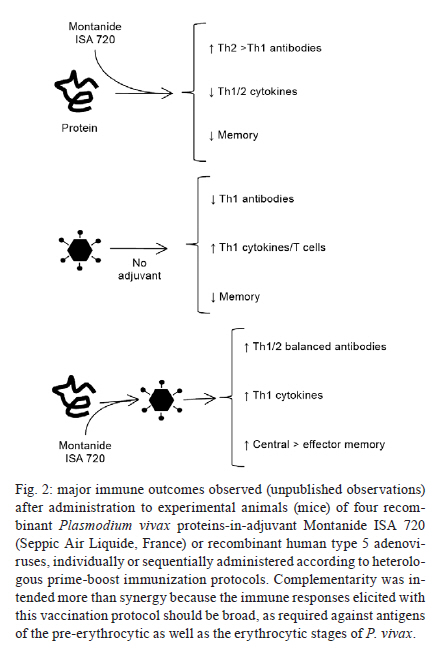
2011 Code Number: oc11157 Abstract The lack of immunogenicity of most malaria antigens and the complex immune responses required for achieving protective immunity against this infectious disease have traditionally hampered the development of an efficient human malaria vaccine. The current boom in development of recombinant viral vectors and their use in prime-boost protocols that result in enhanced immune outcomes have increased the number of malaria vaccine candidates that access pre-clinical and clinical trials. In the frontline, adenoviruses and poxviruses seem to be giving the best immunization results in experimental animals and their mutual combination, or their combination with recombinant proteins (formulated in adjuvants and given in sequence or being given as protein/virus admixtures), has been shown to reach unprecedented levels of anti-malaria immunity that predictably will be somehow reproduced in the human setting. However, all this optimism was previously seen in the malaria vaccine development field without many real applicable results to date. We describe here the current state-of-the-art in the field of recombinant adenovirus research for malaria vaccine development, in particular referring to their use in combination with other immunogens in heterologous prime-boost protocols, while trying to simultaneously show our contributions and point of view on this subject. Key words: malaria vaccines - Plasmodium vivax - Plasmodium falciparum - Adenoviridae - immunization schedule Plasmodium falciparum and Plasmodium vivax cause the vast majority of cases of human malaria worldwide. In mice, Plasmodium yoelii, Plasmodium chabaudi and Plasmodium berghei serve as models for studying different aspects of the human disease - i.e. immunology, pathology or drug/vaccine development - although infection, as well as proteins/antigens generated by these parasites, only partially resembles those of the human species. Even though malaria infection can be treated with drugs, selection of resistant strains and emergence of parasites that can cause severe pathologies are being increasingly documented (Baird 2009, Mita et al. 2009). Enormous efforts are being made to develop a protective anti-malaria vaccine, which will reduce the costs of treatment as well as the social impacts for the regions affected. A commercial malaria vaccine must be feasible, since it has been repeatedly demonstrated that irradiated sporozoites completely protect hosts against the disease (Nussenzweig et al. 1967, Clyde 1990), and antibody transfers from hyperimmune individuals to naÏve subjects are also able to eliminate malaria parasites (McGregor 1964, Clyde 1975). However, the use of irradiated sporozoites for mass vaccination is not strategically achievable and the malaria vaccine that is closer to be translated into a commercial human vaccine, i.e. Glaxo Smithkline's RTS,S/AS01E (Olotu et al. 2011), will probably protect only partially against P. falciparum and nothing against P. vivax. One of the major obstacles for the development of a vaccine against malaria has traditionally been the lack of immunogenicity of the candidate formulations. As an example, researchers believe that protection in RTS,S-vaccinated children is higher when antibody titres against the circumsporozoite protein (CSP) - the plasmodial component of the vaccine - are above 18-40 EU/mL (depending on the analysis model), but neither RTS,S/AS01E nor other RTS,S formulations, are still capable of inducing those titres in all vaccinated kids (Olotu et al. 2011). Amongst the different recombinant vaccine platforms tested during the last decades to address this deficit, viral vectors (Rocha et al. 2004, Li et al. 2007, Limbach & Richie 2009, Hill et al. 2010) represent a significant move forward, for its capacity to improve the potency, functionality and duration of immune responses to refractory vaccine targets, such as CSP, which require complex responses to induce protection. Adenoviruses currently stand as an unavoidable option among the different viruses already tested as vectors for experimental malaria vaccine after repeated demonstrations of their high immunogenicity and capacity of inducing potent T-cell - and to a lesser extent also B-cell - memory immune responses against transgenic products without causing significant pathology (Peiperl et al. 2010). Here, we review the most effective of the use of different adenoviruses vectors along with other vaccine platforms to induce the complex immune responses required to neutralize/eliminate the pre-erythrocytic and asexual erythrocytic stages of the Plasmodium parasite (Table I). Adenoviruses in malaria vaccine development - Most of current's investment in malaria vaccine development is still being made for the development of a P. falciparum vaccine because this parasite is responsible for most fatal cases of the disease, especially in young children in Africa. However, a P. vivax vaccine is also required because vivax malaria is spread worldwide and is gradually becoming more harmful, being responsible for an increasing number of cases of severe and even fatal malaria (Kochar et al. 2005, Tjitra et al. 2008). In both cases, antigens chosen to induce protective immunity in the host must belong to the pre-erythrocytic or the asexual erythrocytic stages of the parasite's life cycle (see the complete cycle somewhere else in this journal issue). Recombinant adenoviruses as vaccine candidates against the pre-erythrocytic stages of malaria - Preventing sporozoites to invade liver cells and/or eliminating infected hepatocytes results in no clinical symptoms of malaria and for P. vivax in no remaining dormant parasites (hypnozoites) that can cause relapses. However, the effector mechanisms underlying protection against Plasmodium pre-erythrocytic stages are complex and clear correlates of immunity are still needed. In addition to components of the innate immune system, such as gamma-delta T-cells, natural killer cells and natural killer T-cells, multifactorial adaptive mechanisms including antibodies, CD8+ and CD4+ T-cells, as well as interferon gamma (IFNγ) have been demonstrated to be key in protection against Plasmodium infection (Doolan & Martinez-Alier 2006, Schwenk & Richie 2011). Nonetheless, the need of inducing strong parasite-specific CD8+ T-cells responses and IFNγ against the pre-erythrocytic forms of Plasmodium seems to be the only prevalent consensus (Ferreira et al. 1986, Schofield et al. 1987). Luckily, these are the most significant traits that result from immunization with most viral vectors and in particular with recombinant adenoviruses (Tatsis et al. 2007) and poxviruses (Harrington et al. 2002). This is because, by nature, most viruses have evolved to be intracellular parasites, and hosts must induce IFNγ and specific CD8+ T-cell responses to promote major histocompatibility complex class I recognition and elimination of infected cells. In the process, not only viral antigens but also heterologous proteins are presented to the immune system, resulting in an outcome that is beneficial to protect against malaria. As previously mentioned, the pre-erythrocytic stage antigen CSP was the first antigen to be vectorized in viruses for immunization. Most viral vectors developed to date have included its recombinant sequence (either complete or the most significant T or B-cell epitopes) in their genomes. Influenza viruses, poxviruses [vaccinia, NYVAC, ALVAC and vaccinia modified ankara virus strain recombinant vector (MVA)], human (HAd) and chimpanzee (AdC) adenoviruses, and others, have documented successful immunization and development of protective CSP-specific CD8+ T-cells responses and IFNγ production. Moreover, the immunological relevance of this antigen is such that even for the first prime-boost immunization protocol ever described (Li et al. 1993), which used influenza and vaccine recombinants, CSP was chosen as model. In the case of recombinant adenoviral vectors, HAd serotype 5 (HAd5V) was initially chosen as the prototype for immunization purposes. The first experimental evidence that HAd5V could be efficiently used as a vector for malaria vaccination was obtained during a study performed by Rodrigues et al. (1997), in which the immunodominant CSP antigen of the P. yoelii sporozoite was expressed in a replication-defective HAd5V vector (Rodrigues et al. 1997). A single immunization with the adenoviral vector elicited unprecedented simultaneous high levels of both cellular and humoral responses against CSP and, in 40% of the mice, complete protection against P. yoelii infection. These encouraging results obtained in animals with the use of HAd5V as vaccine vector - not only for malaria but also for several other diseases - were dampened down when researchers recently suggested that, in humans, the high prevalence of neutralizing antibodies worldwide could neutralize the vector and block vaccination (Pichla-Gollon et al. 2009). In contrast, and despite the initial frustrating outcome reporting lack of protection against human immunodeficiency virus (HIV) infection, Merck's STEP trial showed something very relevant: 75% of human beings were immunized by the three-dose HAd5V-vectored vaccination protocol, irrespectively of their anti-adenovirus immune status before the beginning of the trial (Buchbinder et al. 2008). Since there is no final conclusion on the actual relevance of pre-existing immunity, HAd5V is still in use as a vector, but different strategies are simultaneously being developed to circumvent any possible negative effects that may show up. These include: (i) changing the adenoviral strain to other less frequent human or monkey serotypes (ii) using chimeric capsid-modified adenoviruses or (iii) including the HAd5V vector into heterologous prime-boost protocols that require a maximum of one dose of this vector in combination with others (poxviruses, proteins in different adjuvants etc.). The latter is the alternative supported by our group because does not represent a change in the traits of the immunization vector, while all other options result in changes of the type of target cells infected or in the capacities of the vectors to modulate the immune system, something that has already been demonstrated to change the immunogenic profile of the vaccine candidate (Ophorst et al. 2004, Adams et al. 2011). Regarding the use of the first strategy to avoid pre-existing immunity, researchers at the Dutch company Crucell (Vogels et al. 2003, Ophorst et al. 2004, 2006, Shott et al. 2008) have focused in the use of HAd35V alone or in prime-boost combination with HAd5V, both expressing CSP, and showed that, when combined in a prime-boost protocol, these recombinants adenoviruses induced high CSP-specific cellular and humoral immune responses and, in mice, significantly inhibited liver infection upon a high-dose sporozoite challenge. Moreover, in collaboration with other groups, HAd35V was also tested as vaccine vector to improve immune responses induced by RTS,S/AS01B, a protein-in-adjuvant vaccine. In this study, priming with HAd35V expressing a truncated form of P. falciparum CSP, followed by two RTS,S/AS01B boosters, resulted in IFNγ T-cell re- sponses in a Rhesus monkey model that were significantly increased in both magnitude and duration and kept antibody levels above those induced with three doses of RTS,S/AS01B or with two doses of HAd35V expressing CSP (Stewart et al. 2007). Also at Crucell, Radosevic et al. (2010) are focusing in the use of prime-boost protocols that include HAd26V in combination with HAd35V, HAd5V and/or protein-in-adjuvant formulations, showing in mice and monkeys that some of these combinations induce significant Th1 immune responses against P. falciparum CSP (yeast recombinant) and liver stage antigen 1 (Rodriguez et al. 2009, Radosevic et al. 2010). Future pre-clinical and clinical trials should demonstrate the relative potency of each vector and suggest the final vaccination protocol to be used for vaccination of human beings, but as they own acknowledge, HAd5V should be considered a valid option because of its superior immunogenicity and the fact that only a very small percentage of children under two years (the main target population of a malaria vaccine) carry anti-HAd5V antibodies. Crucell researchers have also tried to develop chimeric viruses (2nd strategy to avoid pre-existing immunity) not for malaria but for HIV infections. In this field, and in collaboration with the group of Dr Daniel Barouch (Nanda et al. 2005), they tested a chimeric HAd35V vector containing HAd5V fibre knobs. This vector displayed immune properties different from the parental serotypes, with a receptor usage and immunogenicity profile similar to that of HAd5V, consequently being more immunogenic than HAd35V. They also reported (Roberts et al. 2006) the use of a second chimeric vector for HIV immunization, based on the HAd5V serotype but with hexon hypervariable regions belonging to the HAd48V serotype. This vector induced in mice cellular immune responses against HIV gag protein comparable to those induced by HAd5V vector without significant neutralization by pre-existing immunity. However, none of these vectors were yet reported as malaria vaccine candidates yet, so we cannot evaluate their properties at this time. Working in malaria vaccine development, Shiratsuchi et al. (2010) constructed a novel HAd5V recombinant that displayed the main B-cell epitope of P. yoelii CSP within the adenovirus hexon and/or fibre, while expressing the full-length P. yoelii CSP within infected cells, aiming at the simultaneous induction of potent B-cell and T-cell responses. Viruses with the hexon hypervariable region 1 replaced with the CSP B epitope induced not only the highest humoral response but also significant protection against a subsequent malaria challenge and, most importantly, circumvented neutralization by pre-existing anti-HAd5V antibodies. For malaria prevention also, Palma et al. (2011) constructed a recombinant HAd5V expressing two different B cell epitopes of P. falciparum CSP within the hexon protein. Different from all others studies cited here, these two recombinant vectors were replicative. Results show that CSP epitopes displayed on adenoviruses particles could elicit neutralizing antibodies, even when tested in mice, animals in which adenoviruses neither replicated nor expressed late proteins (including hexon). The third strategy mentioned above, hence, the use of heterologous prime-boost protocols that involve administration of a single dose of HAd5V recombinants, has been our choice for the development of an anti-malaria vaccine. In the case of Plasmodium pre-erythrocytic antigens, and based in previous works from Dr Ruth and Victor Nussenzweig's laboratories at New York University (Li et al. 1993) that resulted in the description of the first prime-boost immunization protocols, we showed a decade ago (Bruna-Romero et al. 2001) that immune responses generated by the CSP-recombinant HAd5V vector could be further improved by the sequential administration of a poxviral recombinant vector (VacPyCS), a western reserve vaccinia virus recombinant expressing the same antigen, as a booster. This heterologous prime/boost protocol was able to protect 100% of experimental animals against murine malaria after a challenge with live P. yoelii sporozoites. Currently, our experiments with P. vivax antigens are being designed to even further increase the immunogenicity of CSP. For this, CSP has been included within the hepatitis B virus core antigen (HBcAg) to form a viral-like particle or, alternatively, fused to the herpes simplex virus I (HSV-1) glycoprotein D carrier protein (gD). Both chimeras are being administered either in their purified forms or expressed in loco by means of recombinant adenoviruses. The HBcAg spontaneously assembles into virus-like particle composed of 180-240 monomers, that are highly immunogenic for B-cell epitopes inserted into the immunodominant loop region located at the tip of the surface "spikes" on particles mounted with HBcAg, as well as for the T-cell epitopes placed at the C-terminus of the same molecule (Francis et al. 1990, Bottcher et al. 1997, Birkett et al. 2002). HSV-1 gD binds to the herpes virus entry mediator and stimulates antigen presentation by dendritic cells and inhibition of Tregs (Murphy et al. 2006, DiMenna et al. 2010). Based on our current results, it is our hypothesis that these recombinant imunogens, i.e. the chimeric HBcAg and gD pure proteins, given alone or in combination with adenoviruses that are able to express them in infected host cells, will generate simultaneous, potent and long-lasting CSP-specific cellular and humoral immune responses (hypothesis explained in Fig. 1). Another antigen that has also been used in vaccine formulations against the pre-erythrocytic stages of Plasmodium is the thrombospondin-related anonymous protein (TRAP)/sporozoite surface protein 2. The group of Dr Adrian Hill and colleagues at Oxford University have been using this antigen in a proprietary design, known as the multiepitope (ME)-TRAP (Moorthy et al. 2003) a transgene that contains P. falciparum TRAP fused in frame to a ME string of B, CD4+ and CD8+ epitopes from CSP and others. This antigen was initially used as a DNA vaccine, further complemented by the addition of a poxviral MVA vector to boost responses. More recently, adenoviral vectors AdC63 and AdC9 have also been included in the immunization routines (Capone et al. 2010, Reyes-Sandoval et al. 2010) and authors claim that, by using these recombinant virus combinations, they efficiently induce potent Th1 polyfunctional protective T cells. Recombinant adenoviruses as vaccine candidates against the erythrocytic stages of malaria - In principle, since human leukocyte antigen class I or II molecules are absent from the surface of the parasite or the infected red blood cell, the focus in an asexual blood-stage malaria vaccine is the induction of antibodies against intra-erythrocytic and free blood forms of Plasmodium (Fowkes et al. 2010). These antibodies should neutralize free merozoites, stop further infection of red blood cells and bring to an end subsequent multiplication of the parasite. This type of vaccines may not be sterilizing, but greatly reduce/eliminate symptoms and pathology. This simplistic view is, however, much more complex when the detailed immune mechanisms involved are studied. Thus, to induce potent and efficient antibody responses and memory B-cells, T-cell help and cytokine production are still required and some authors (McCall & Sauerwein 2010) have described a direct relation between protection and specific T-cell induction against blood-stage parasite antigens. Thus, at the end of the road, vaccination must not only focus on antibodies but also on generation of specific T-cells. The main blood-stage candidates currently in study for development of a recombinant adenoviral vaccine against Plasmodium are: apical membrane antigen 1 (AMA-1), merozoite surface protein 1 (MSP-1) and Duffy-binding protein (DBP), the latest being of particularly importance to prevent P. vivax infection. AMA-1 is a type I integral membrane protein found in all species of Plasmodium. This protein is frequently described as a blood-stage malaria vaccine candidate, but some studies suggest that AMA-1 has a dual role during Plasmodium infection, acting both in the pre-erythrocytic (Silvie et al. 2004) and erythrocytic (Triglia et al. 2000) stages, and antibodies against AMA-1 have been demonstrated to block sporozoite invasion of the hepatocytes and also prevent merozoite invasion of red blood cells. In animal models, antibodies directed against AMA-1 induce protection against Plasmodium challenges, but clinical trials using AMA-1 protein administered in different adjuvants showed limited immunogenicity in humans (Saul et al. 2005, Hu et al. 2008, Malkin et al. 2008, Roestenberg et al. 2008, Pierce et al. 2010). To improve the immunogenicity of AMA-1 and strongly enhance the cellular immune responses that could stop the development of both pre-erythrocytic as well as erythrocytic forms, recombinant viral vectors are being used together or as an alternative to protein-based vaccines (Draper et al. 2010). MSP-1 is synthesized from the onset of schizogony as a 195 kDa precursor that undergoes a series of proteolytic cleavages during invasion of host erythrocytes. After the first proteolytic cleavage only a fragment of 42 kDa (MSP-142) remains attached to the membrane. This fragment is then cleaved again generating two others, MSP-133 (soluble) and MSP-119 (glycosylphosphatidylinisotol-anchored) (Kadekoppala & Holder 2010). MSP-142 and MSP-119 are leading vaccine candidates since there is evidence of protection mediated by antibodies against these molecules (John et al. 2004, Stanisic et al. 2009). However, to date, when tested as proteins-in-adjuvants in combination with different immune stimulants, they did not protect immunized humans (Malkin et al. 2007, Ogutu et al. 2009). DBP is a 140 kDa protein that serves as a P. vivax ligand for a chemokine receptor present on the surface of the erythocytes, the Duffy antigen/receptor for chemokines, also known as the Duffy blood group. Since P. vivax is highly dependent on interaction with the Duffy blood group to invade human erythrocytes, Duffy-negative humans are mostly (although it has been shown that not in all cases) resistant to invasion by P. vivax and antibodies against this molecule can prevent attachment and/or invasion of erythrocytes (Miller 1976). Antibodies against DBP have been associated with protection in humans (King et al. 2008) and in vaccinated animals inhibited erythrocyte adherence in vitro (Michon et al. 2000). Regarding the development of adenoviral vaccine candidates encoding these three antigens, several reports have been published recently. Bruder et al. (2010) tested in rabbits the impact of glycosylation and cellular localization of P. falciparum AMA-1 and MSP-142 expressed by recombinant HAd5V vectors on the immune responses detected after immunization. Their results show that when secreted to the cell surface, both antigens induce significantly higher levels of antibodies and T cells against both antigens and that glycosylation does not interfere with functionality. In this study, humoral immune responses induced by the HAd5V adenovectors elicited 99% and 60% inhibition of parasite growth, respectively for each antigen. To achieve broad, polyfunctional and potent immune responses in animal models Draper et al. (2010) used AdC63 and MVA, both expressing P. falciparum AMA-1, and also AMA-1 protein/virus admixtures for heterologous prime-boost immunizations of Rhesus monkeys. Authors show that sequential immunization with adeno and poxviral vectors induce strong and long-lasting, polyfunctional [tumor necrosis factor alpha (TNFα), IFNγ and interleukin (IL)2-secreting] CD8+ and CD4+ T-cell responses that exhibit a central memory-like phenotype. Immunizations with AdC63-MVA-protein or AdC63-protein-protein regimens, regardless of the adjuvant used, induced B-cells and high titre functional IgG responses that inhibited the growth of P. falciparum in vitro. The authors did not test, however, the possibility of a first dose of protein-in-adjuvant followed by an adenoviral vector booster dose, which in our hands has been the most efficient immunization protocol amongst all studied to date, not only for P. vivax AMA-1, but also for the other two blood-stage antigens we routinely use for immunization, i.e. MSP-1 and DBP. Goodman et al. (2010) studied the immunogenicity of two allelic forms of P. falciparum MSP-142 when administered as a HAd5V, distinct serotypes of AdC or as a recombinant MVA vector. Authors suggest that replacing HAd5V with simian adenoviral vector compromised neither antibody nor T cell responses in mice, leading to antibody titres similar to those obtained by three immunizations with protein in Freund's adjuvant. Further comparisons in other animal models and in clinical trials are expected, which will help to validate these results. In a similar work, comparing immunization protocols based in AdC, MVA and protein in adjuvant, Douglas et al. (2010) highlighted the importance of testing tailored protocols to obtain the immune responses pretended. They showed that protocols containing AdC63 and MVA induced maximal CD8+ T cell responses while maximal IgG responses were elicited by protocols containing AdC63 and protein. They also showed the feasibility of using an admixture of the viral vectors with proteins in order to simplify the immunization regiments, since two-shot immunizations with admixed antigens elicited similar responses than three-shot immunizations. All reports regarding recombinant adenoviruses mentioned up to here have used P. falciparum antigens. Our research group is however very interested in developing a vaccine against P. vivax. We have pursued the idea that a heterologous prime-boost immunization regime that includes administration of an antigen as an adequate protein-in-adjuvant (Montanide ISA720 preferred) formulation, followed by the administration of an adenovirus vector, could be one of the best approaches to achieve longer-lasting simultaneous B and T immune responses against P. vivax. Immune traits of each of these two immunogens when given separately or in combination are depicted in Figs 1 and 2 and led us to propose this hypothesis. In a proof of concept of this methodology we are using the three main blood-stage antigens described here. Thus, we have expressed in Escherichia coli and in eukaryotic cells (by means of 3 independent recombinant HAd5 viruses) the extracellular domains of AMA-1 (Rodrigues et al. 2005), a chimeric form of MSP-119, named MSP-119-PADRE, in which the universal T Pan-Allelic DR epitope is fused to the C-terminus of P. vivax MSP-119 sequence (Cunha et al. 2001) and two distinct polymorphisms of the first two extracellular domains of DBP (Sousa et al. 2006). Our results (unpublished observations) indicate that administration of P. vivax proteins in the synthetic adjuvant Montanide ISA 720 (Lawrence et al. 1997) followed by administration of HAd5V vectors expressing equivalent antigens elicit long-term B and T cell central as well as effector memory responses, with high levels of IgG1 and IgG2a antibodies. Monkey experiments are currently on their way to define the safety, immunogenicity and protective efficacy of the prime-boost protocol and the P. vivax antigens. Concluding remarks - Adenoviral-vectored vaccines have demonstrated its outstanding value as components of prime-boost protocols for induction of immune responses, in particular polyfunctional cell-mediated (IFNγ/IL2/TNFα-producing CD8+ T cells) responses, against malaria. In combination with other (mostly viral) platforms, adenoviruses have become an irreplaceable tool to induce immunity against the pre-erythrocytic stages of malaria. Administration of this vaccine platform along with protein-based vaccines may also result in improved levels of Plasmodium-specific antibodies and CD4+ T cells, of extreme interest when fighting the blood-stages of the parasite. Combinations of different viruses (at present the most efficient would be adenoviruses and poxviruses) and/or protein-in-adjuvants in "tailored" immunization protocols, which can be different depending on the pre-erythrocytic or asexual blood-stage antigens used, could end-up solving the immunogenicity problems that malaria vaccine candidates have faced to date. However, still several pre-clinical and clinical trials will have to be performed to define the best adenoviral serotypes and immunization protocols to be used, before firm vaccine candidates may enter industrial pipelines. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11157t1.jpg] [oc11157f1.jpg] [oc11157f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}