 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 212-217 Original Article Can Wolbachia be used to control malaria? Thomas WalkerI; Luciano Andrade MoreiraII, + ISchool
of Biological Sciences, The University of Queensland, Brisbane QLD, Australia Received 10 March
2011 Code Number: oc11159 Abstract Malaria is a
mosquito-borne infectious disease caused by Plasmodium parasites transmitted
by the infectious bite of Anopheles mosquitoes. Vector control of malaria has
predominantly focused on targeting the adult mosquito through insecticides and
bed nets. However, current vector control methods are often not sustainable
for long periods so alternative methods are needed. A novel biocontrol approach
for mosquito-borne diseases has recently been proposed, it uses maternally inherited
endosymbiotic Wolbachia bacteria transinfected into mosquitoes in order to interfere
with pathogen transmission. Transinfected Wolbachia strains in Aedes aegypti
mosquitoes, the primary vector of dengue fever, directly inhibit pathogen replication,
including Plasmodium gallinaceum, and also affect mosquito reproduction
to allow Wolbachia to spread through mosquito populations. In addition, transient
Wolbachia infections in Anopheles gambiae significantly reduce Plasmodium
levels. Here we review the prospects of using a Wolbachia-based approach to
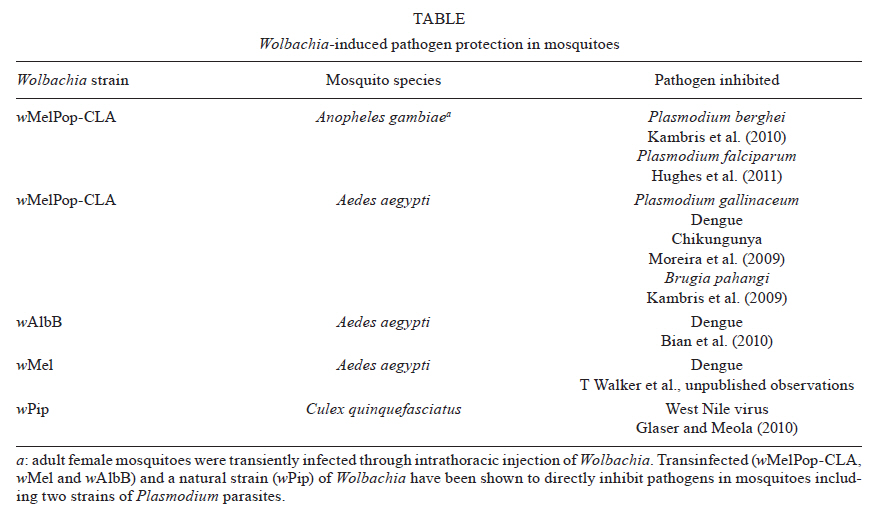
reduce human malaria transmission through transinfection of Anopheles mosquitoes. Wolbachia's phenotypic effects in insect hosts - Wolbachia are Gram-negative, intracellular, endosymbiotic bacteria that manipulate host reproduction to enhance their vertical transmission (Sinkins et al. 1997). Wolbachia bacteria were first reported within the reproductive tissues of Culex pipiens mosquitoes by Hertig and Wolbach in 1924 and the species was named Wolbachia pipientis (Werren 1997). Infections are extremely widespread within arthropods (Werren et al. 1995, O'Neill et al. 1997, Jeyaprakash & Hoy 2000) and Wolbachia are present in numerous mosquito genera including Aedes, Culex, Coquillettidia and Mansonia (Kittayapong et al. 2000, Ricci et al. 2002, Dean & Dobson 2004, Tsai et al. 2006). Wolbachia are maternally transmitted through the egg cytoplasm and are responsible for several reproductive disorders in their insect hosts such as cytoplasmic incompatibility (CI) (Yen & Barr 1971), parthenogenesis (Stouthamer et al. 1999), feminization (Rousset et al. 1992) and male killing (Hurst et al. 2000). CI results in the generation of unviable offspring when an uninfected female mates with a Wolbachia-infected male (McGraw et al. 2001). In contrast, Wolbachia-infected females can produce viable progeny when they mate with both infected and uninfected males resulting in a selective reproductive advantage over uninfected females (Hoffmann & Turelli 1997). This CI phenotype, shown in Figure, is induced by Wolbachia in mosquito species and allows the maternally transmitted Wolbachia to efficiently invade host populations without being infectious or moving horizontally between individuals (Hoffmann & Turelli 1997). Although Wolbachia usually form non-virulent associations with their hosts (Sinkins et al. 1997), a virulent strain of Wolbachia, wMelPop, has been described that reduces the adult life span of its natural fruit fly host Drosophila melanogaster (Min & Benzer 1997). This Wolbachia infection appears to be quiescent in the developing immature stages of the fruit fly but enters into a stage of massive over replication once adult flies emerge. The life-shortening phenotype induced by the wMelPop strain was predicted to have potential use in the control of mosquito-borne diseases. As most pathogens need to develop within the mosquito vector for 8-21 days post-infection, removal of older mosquitoes from the population, which are responsible for the majority of transmission, may result in substantial reductions in disease transmission (Sinkins & O'Neill 2000, Brownstein et al. 2003, Rasgon et al. 2003, Cook et al. 2008). However, the major vectors of human diseases such as Anopheles species (malaria) and Aedes aegypti (dengue) do not harbour natural Wolbachia infections. Therefore, the primary aim of using Wolbachia for human disease control is the stable transinfection of the bacteria into medically important mosquito vectors. Wolbachia transinfection of Ae. aegypti mosquitoes - Successful transinfection of Wolbachia between distantly related insect species is dependent on the ability of the Wolbachia strain to adapt to new intracellular environments (Braig et al. 1994, Xi et al. 2005a). The wAlbB strain of Wolbachia was successfully established in Ae. aegypti using embryo cytoplasm transfer from closely related Aedes albopictus mosquitoes (Xi et al. 2005b). However, to facilitate the transfer of Wolbachia strains from more distantly related insect hosts, mosquito cell line adaptation appears to be critical for transinfection success. In order to facilitate the transfer of wMelPop from D. melanogaster fruit flies into Ae. aegypti mosquitoes, the bacteria was first transferred into mosquito cell lines to allow adaptation to the mosquito intracellular environment (McMeniman et al. 2008). After continuous serial passage in mosquito cell culture for over three years, the mosquito cell line-adapted Wolbachia strain, wMelPop-CLA, was stably introduced into Ae. aegypti using embryo microinjection (McMeniman et al. 2009). Two Wolbachia-infected lines were generated after a period of experimental selection in early generations and both lines remain highly infected four years after establishment. The wMelPop-CLA strain results in approximately 50% reduction in the adult lifespan of Ae. aegypti mosquitoes (McMeniman et al. 2009, Yeap et al. 2011). This reduction in adult lifespan of female mosquitoes is predicted to result in a significant reduction in dengue transmission, if the capacity for life-shortening under laboratory conditions can be translated into field settings. Mosquito age is a critical factor for pathogen transmission (Dye 1992) as pathogens such as Plasmodium parasites undergoes an extrinsic incubation period (EIP) within the mosquito. The EIP is the time required from the ingestion of the pathogen until it is transmitted to the next vertebrate host. The speed of Plasmodium development within Anopheles mosquitoes depends on host, parasite and environmental factors such as temperature (Paaijmans et al. 2009). A typical incubation period of 10-14 days is relatively long compared to the longevity of adult mosquitoes (Charlwood et al. 1997, Killeen et al. 2000). In addition, there is typically at least two days from adult eclosion until adult female mosquitoes take their first bloodmeal. Adult mosquitoes also experience a high daily mortality rate resulting in only a small percentage of the total population actually surviving long enough to transmit malaria (Brownstein et al. 2003). Therefore, a reduction in the daily survival rates is likely to remove a large proportion of the mosquito population capable of transmitting malaria. Currently the most effective malaria vector control strategies, including indoor residual insecticide sprays and long-lasting insecticide-treated net, reduce the daily survival rates of Anopheles mosquitoes (Enayati & Hemingway 2010). Reducing the survival of mosquitoes leads to an approximately exponential decline in transmission intensity (Bellan 2010). Consequently, transinfection of life-shortening strains of Wolbachia into Anopheles would be predicted to significantly impact malaria transmission. Wolbachia-induced pathogen interference - In Drosophila, the wMelPop and closely related Wolbachia strains have the capability of protecting against RNA virus infection by delaying the mortality of flies infected with a range of pathogenic viruses (Hedges et al. 2008, Teixeira et al. 2008). Wolbachia strains also provide significant pathogen protection in mosquitoes (Table). Infection of the wMelPop-CLA strain in Ae. aegypti also provides direct resistance against dengue infection (Moreira et al. 2009). When dengue serotype-2 (DENV-2) was introduced into wMelPop-CLA infected Ae. aegypti mosquitoes by either intrathoracic injection or oral feeding, an almost complete protection against viral infection was observed. The ability of wMelPop-CLA to interfere with viral replication also appears to occur with other arboviruses transmitted by Ae. aegypti, with similar virus interference effects for Chikungunya virus in oral feeding experiments of wMelPop-CLA infected Ae. aegypti (Moreira et al. 2009). There is also further evidence that the wMelPop-CLA strain provides protection against both filarial nematodes (Kambris et al. 2009) and avian malaria parasites (Moreira et al. 2009) suggesting some Wolbachia strains may inhibit a broad range of human pathogens. As viral interference is not ubiquitous among Wolbachia strains (Moreira et al. 2009, Osborne et al. 2009) the mechanisms behind the ability of Wolbachia to provide resistance against pathogens are unknown. Although immune effector genes are upregulated in wMelPop-CLA infected Ae. aegypti mosquitoes, key components of the currently accepted signalling pathways for these effectors do not appear to be transcriptionally modulated by Wolbachia (Moreira et al. 2009, Kambris et al. 2010). Previous studies also revealed that some genes from the Toll, Imd and Jak-STAT pathways, implicated in the control of RNA virus infection in insects (Huszar & Imler 2008) are differentially regulated in Ae. aegypti mosquitoes infected with dengue (Xi et al. 2008). The ability of the wMelPop-CLA strain to provide dengue virus protection may also be dependent on competition for essential host cell components, as DENV-2 infection was only observed in cells of wMelPop-CLA infected mosquitoes that did not harbour Wolbachia (Moreira et al. 2009). Interestingly, cholesterol is a key fatty acid that is obtained from the insect host cell by both Wolbachia (Lin & Rikihisa 2003, Wu et al. 2004) and arboviruses (Lu et al. 1999, Mackenzie et al. 2007). In addition, Plasmodium is dependent on host lipids in the mosquito stage (Atella et al. 2009), suggesting cholesterol could be a critical host nutrient required by both Wolbachia and mosquito-borne pathogens. The density and tissue distribution of Wolbachia infections in insect hosts may be an important determinant of their ability to interfere with pathogens. Wolbachia strains that provide protection in Drosophila simulans are closely related to the wMelPop strain in D. melanogaster and are found at comparatively high densities in flies (Osborne et al. 2009). The closely-related non-virulent wMel strain also provides significant protection against DENV-2 in transinfected Ae. aegypti mosquitoes resulting in complete blockage of dengue transmission under experimental conditions (unpublished observations). Interestingly, strains of Wolbachia that naturally reside in mosquitoes show no or very limited capability for virus protection. Ae. albopictus mosquitoes are infected with non-virulent wAlbA and wAlbB strains of Wolbachia (Sinkins et al. 1995) yet are competent vectors of dengue virus (Kyle & Harris 2008). Similarly, Armigeres subaltatus mosquitoes are infected with a Wolbachia strain but no evidence is seen for interference with Japanese encephalitis virus (Tsai et al. 2006). Aedes fluviatillis mosquitoes are infected with a strain of Wolbachia, named wFlu, despite being competent laboratory vectors of Plasmodium gallinaceum (Moreira et al. 2009). Recently the native wPip strain of Wolbachia in Culex quinquefasciatus was shown to have some protective effect against West Nile virus (Glaser & Meola 2010). However, this effect was much less pronounced when compared to the effects on dengue virus for transinfected fruit fly Wolbachia strains in Ae. aegypti (T Walker et al., unpublished observations). Overall it appears that the ability of Wolbachia to generate pathogen interference is likely to be restricted to Wolbachia strains that grow to high densities and have a wide tissue tropism in their insect host. Wolbachia transinfection of Anopheles mosquitoes - The use of Wolbachia for malaria control will require a stable infection that is transmitted vertically to offspring, as occurs with fruit fly Wolbachia strains transinfected into Ae. aegypti mosquitoes (McMeniman et al. 2009, T Walker et al., unpublished observations). Natural Wolbachia infections had never previously been detected in any species of Anopheles (Curtis & Sinkins 1998, Kittayapong et al. 2000, Ricci et al. 2002, Tsai et al. 2004, Slotman et al. 2005). However, Wolbachia-positive individuals have recently been found in some species of Anopheles from the Amazon Region (RA Passos & WP Tadei, personal communication). Further studies on this finding can greatly open the potential use of this methodology for malaria control. The challenging nature of mosquito microinjection has hampered the progress in transferring Wolbachia between mosquito species. The transfer of Wolbachia into mosquito embryos is intrinsically more difficult than transfer into Drosophila, since mosquito embryos are less amenable to inoculation, especially Anopheles embryos. Factors such as the age at the time of injection and desiccation level are especially critical for both injectability and survival of Anopheles embryos. Using Wolbachia-infected mosquito cell lines, the generation of a stably infected line was not been possible despite microinjection of approximately 10,000 embryos of the malaria vectors Anopheles stephensi and Anopheles farauti (unpublished observations). Wolbachia strains can be maintained in vitro in immunocompetent Anopheles gambiae cell lines (Rasgon et al. 2006, McMeniman et al. 2008), suggesting there is no intrinsic genetic mechanism preventing the infection of Anopheles cells with Wolbachia. In addition, when the wMelPop strain was injected into the hemolymph of An. gambiae adult females, the bacteria were able to survive and replicate in somatic tissues (Jin et al. 2009). However, the infection was not present in germline tissue (ovaries) so a stable transinfection could not be established using this methodology. The reason why Wolbachia does neither naturally infect Anopheles species nor form a stable infection is unknown. The ability to generate transient somatic infections through injection of adult mosquitoes (Jin et al. 2009, Kambris et al. 2010, Hughes et al. 2011) suggests the possibility that Wolbachia may be unable to colonize the germline tissue (ovaries). Successful transinfection requires infection of the germline prior to pole-cell formation in the pre-blastoderm stage of embryo development. Maternal transmission of Wolbachia to resulting progeny is dependent on establishing infections in the ovaries of adult females. Ultimately this barrier to germline infection must be overcome to establish stably infected lines that could be deployed for malaria vector control strategies. A recent additional study shows Plasmodium falciparum development in An. gambiae is surpressed by transient somatic infections of wMelPop-CLA (Hughes et al. 2011). Wolbachia's effect on Plasmodium - Several studies present evidence that Wolbachia is likely to provide some protection against human malaria Plasmodium parasites if stable transinfection of Anopheles is achieved. The effect of the wMelPop-CLA strain on P. gallinaceum was tested as this species of malaria parasite is known to be able to infect Ae. aegypti mosquitoes in the laboratory. The P. gallinaceum oocyst load was reduced by 67-88% for wMelPop-CLA infected Ae. aegypti mosquitoes compared to Wolbachia-uninfected mosquitoes seven days after feeding on an infected chicken (Moreira et al. 2009). In An. gambiae females transiently infected with wMelPop using adult injection, mean Plasmodium berghei levels were reduced by 75-84% (Kambris et al. 2010). Although this combination of vector/parasite does not occur in nature, these results do highlight the ability of Wolbachia to significantly reduce the levels of malaria parasites in Anopheles mosquitoes. Wolbachia invasion of wild Anopheles mosquito populations - An introduced Wolbachia infection in Anopheles mosquitoes would require induction of the CI phenotype and high rates of maternal transmission to successfully invade wild populations. The wMelPop and wMel strains induce CI in transinfected Ae. aegypti mosquitoes (Mc-Meniman et al. 2009, T Walker et al., unpublished observations), suggesting this phenotype would be induced in stably infected Anopheles lines. The maternal transmission rate of wMelPop-CLA in Ae. aegypti is estimated to be above 99% in laboratory lines (McMeniman et al. 2009), suggesting a high infection frequency could be achieved in transinfected Anopheles lines. The potential of Wolbachia to rapidly spread through insect populations under the action of CI was spatially described by Turelli and Hoffmann (1991) in wild D. simulans populations in California with the wRi strain rapidly spreading at a rate of 100 km per year. This rapid spread of Wolbachia through wild Drosophila populations was described as a "Bartonian wave" (Turelli & Hoffmann 1991). The ability of any Wolbachia strain to successfully invade wild Anopheles mosquito populations will depend on an unstable threshold infection level. This unstable point depends on the negative selection imposed by fitness costs of Wolbachia infection and positive selection associated with CI induction (Hoffmann & Turelli 1997, Turelli 2010). If the threshold infection frequency is reached through introduction at an initial prevalence greater than the unstable equilibrium value, the Wolbachia infection is expected to spread to fixation over subsequent generations (Hoffmann & Turelli 1997, Turelli 2010). Any fitness costs imposed by Wolbachia will raise the unstable point slowing the spread of infection through the "Bartonian wave" of invasion. The likely fitness costs associated with the stable introduction of the wMelPop-CLA strain into Anopheles will make this strain relatively weak at spreading into mosquito populations. Alternative Wolbachia strains such as wMel that are predicted to impose less of a fitness cost to transinfected Anopheles mosquitoes are likely to be more successful at invasion of wild mosquito populations. The minimal fitness costs of Wolbachia infection are critical given the importance of mosquito fitness on the ability of released mosquitoes to compete with wild populations. A major advantage of a Wolbachia-based biocontrol approach for malaria is that CI acts as a self-spreading mechanism for Wolbachia to rapidly invade populations from the release of relatively small numbers of individuals. The predicted direct inhibition of human Plasmodium parasites by Wolbachia may also augment CI as a broad based mechanism for population invasion to provide a positive fitness benefit to Anopheles mosquitoes carrying Wolbachia. This may overlay the traditional "Bartonian" view of CI based invasion dynamics and provide an additional driving force for Wolbachia depending on the extent of fitness advantages conferred to field populations. Ultimately the use of Wolbachia for malaria control will require stably infected lines of major malaria vectors such as An. gambiae s.l. (Africa), An. stephensi (India) and An. darlingi (Central and South America) and a comprehensive assessment of the protective effect against human malaria parasites such as P. falciparum and P. vivax. The applied use of Wolbachia for malaria control would also require significant characterization of Wolbachia's phenotypic effects in diverse genetic background of these Anopheles vector species. In reality, widespread control of malaria using Wolbachia-based methods is not likely achievable. For example, the difficulties of colonizing An. darlingi (and therefore transinfecting this species with Wolbachia) would prevent the applied use of Wolbachia for control of malaria in parts of the Amazonian Region. In that case, transinfection of colonisable species such as Anopheles aquasalis (Da Silva et al. 2006) would provide applicability in areas where this species has vectorial importance. Lastly, one has to be aware that the complexity of malaria vector populations (Lanzaro et al. 1998, Donnelly et al. 2002) would be a major complicating factor in the applied use of Wolbachia for malaria control. However, this novel approach may provide an effective mechanism of malaria control in some malaria endemic areas in which a single, vector species is present. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11159t1.jpg] [oc11159f1.jpg] |
| |||||||||

{kind=link}
{kind=link}