 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 106, Special Issue, pp. 223-238 Original Article Malaria vector species in Colombia - a review James Montoya-LermaI, +; Yezid A SolarteII, III; Gloria Isabel Giraldo-CalderónI; Martha L QuiñonesIV; Freddy Ruiz-LópezV; Richard C WilkersonV; Ranulfo GonzálezI IBiology
Department Received 1 April
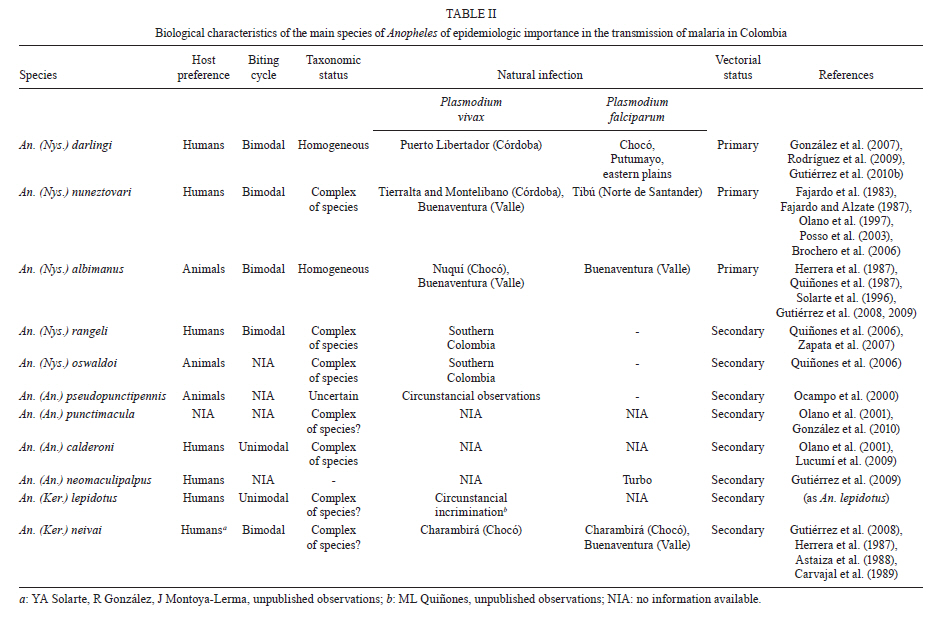
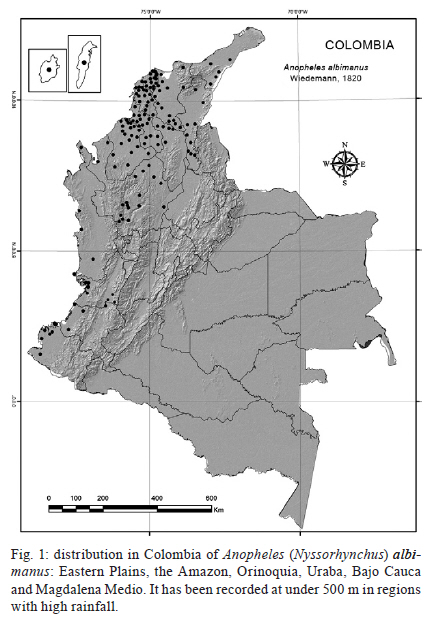
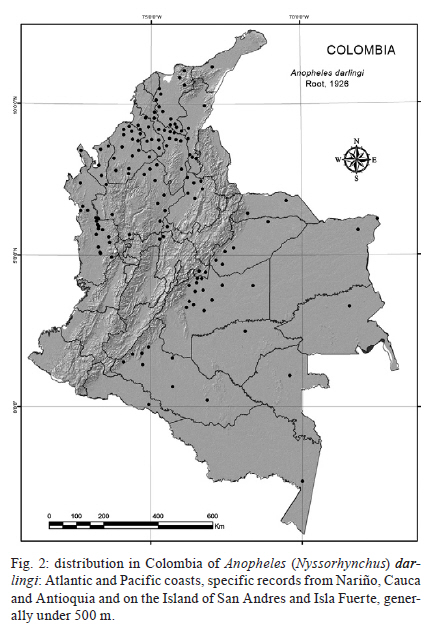
2011 Code Number: oc11163 Abstract Here we present a comprehensive review of the literature on the vectorial importance of the major Anopheles malaria vectors in Colombia. We provide basic information on the geographical distribution, altitudinal range, immature habitats, adult behaviour, feeding preferences and anthropophily, endophily and infectivity rates. We additionally review information on the life cycle, longevity and population fluctuation of Colombian Anopheles species. Emphasis was placed on the primary vectors that have been epidemiologically incriminated in malaria transmission: Anopheles darlingi, Anopheles albimanus and Anopheles nuneztovari. The role of a selection of local, regional or secondary vectors (e.g., Anopheles pseudopunctipennis and Anopheles neivai) is also discussed. We highlight the importance of combining biological, morphological and molecular data for the correct taxonomical determination of a given species, particularly for members of the species complexes. We likewise emphasise the importance of studying the bionomics of primary and secondary vectors along with an examination of the local conditions affecting the transmission of malaria. The presence and spread of the major vectors and the emergence of secondary species capable of transmitting human Plasmodia are of great interest. When selecting control measures, the anopheline diversity in the region must be considered. Variation in macroclimate conditions over a species' geographical range must be well understood and targeted to plan effective control measures based on the population dynamics of the local Anopheles species. Key words: Anopheles - Colombia - review Recent reports on malaria reveal that in the Americas malarial morbidity decreased by 32%, from 1.1 million cases in 2000 to 775,000 cases in 2007 (WHO 2008, Sinka et al. 2010). Morbidity in Colombia specifically followed the same trend, but with less intensity. For 2000 and 2007, the morbidity figures were 144,432 and 128,462, respectively, with a decrease of 11%, contrasting with data from neighbouring countries with the exception of Venezuela (WHO 2010). Several factors have limited the effort to control human malaria in Colombia, including the lack of adequate knowledge of vector control activities and the misapplication of insecticide (usually due to inadequate frequency) as well as to the drug resistance of some Plasmodium populations to available treatments (Olano et al. 1997, Carmona-Fonseca 2003, 2004). These factors allow the malaria parasites to maintain its virulence and its importance as a human pathogen. Additionally, socio-economic, political and public safety conditions complicate the surveillance of patients and vectors (Beyrer et al. 2006). The situation has become more critical with the development of urban transmission in some regions (Méndez & Carrasquilla 1995, Porras et al. 2001, Ochoa & Osorio 2006), the presence of new species (González et al. 2010, Ruiz et al. 2010) and the presence of species complexes (Brochero et al. 2010, Gutiérrez et al. 2010a).The understanding of mosquito vector biology and ecology derived from field research and laboratory studies was directed towards understanding the bionomics of Anopheles vectors and was useful as a determinant for designing adequate measures to control malaria (Gabaldón 1983) and to set up a global strategic framework for integrated vector management (IVM) (WHO 2004). Although the use of pesticides facilitated the suppression of malaria in many areas and its control in others (Enserink 2002), it led to false expectations of their efficacy and resulted in their misuse and, in some cases, the development of pesticide resistance (Fonseca-González et al. 2010). At present, to fulfil the goals of the Global Malaria Action Plan (RBM 2008), the Colombian Social Protection Ministry has undertaken the challenge of defining and implementing a broad-based control strategy. The prospects for releasing refractive, transgenic malaria vector species are distant due to economical constraints and limited knowledge of the bionomics of the major vector species. Therefore, a sustained, integrated vector control strategy (Rojas et al. 2001, Snow et al. 2008) represents the most feasible and economical alternative. Effective vector control has been limited by the insufficiency of information on the species of Anopheles mosquitoes that serve as malaria vectors; some of them are now recognised as species complexes, which include vector and non-vector species with different biologies, distributions and responses to control measures (Brochero & Quiñones 2008). Additionally, uncertainties regarding the impact of anthropogenic environmental changes on the dynamics of malaria parasite transmission and the limited availability of effective tools for IVM have impacted the success of vector control. The goal of the Centro Latino Americano de Investigación en Malaria with respect to vector biology is to address the major gaps in our understanding of the ecology, behaviour, vector potential and control of Anopheles malaria vectors to guide the development and implementation of more effective IVM strategies in Colombia and Latin America. Recently, Sinka et al. (2010) developed a series of predictive species range maps by compiling contemporary data on the occurrence of the dominant malaria vector species and the environmental and climatic variables relevant to anopheline ecology. Although it was tempting to use the same approach here, the absence of reliable databases for the dominant (sensu Sinka et al. 2010) Colombian vector species hindered the use of this approach. Therefore, for this review, we opted for the conservative designation of the primary and secondary vector species. We review and summarise the existing information regarding the bio-ecological characteristics of the major malaria vector species in Colombia. The geographical distributions, breeding habitats, feeding habits, host-seeking activities and resting patterns of the primary and secondary vectors are analysed and discussed. Characteristics of vectors - According to Donnelly et al. (2002), an anopheline species is a primary vector if it possessed the following ecological and physiological characteristics: (i) wide geographic distribution, (ii) high, although frequently seasonal, local abundance, (iii) significant dispersion and colonisation capacities, (iv) adaptations for exploiting different man-made environments and commodities (e.g., buildings, cultivated areas and vehicles), (v) haematophagy, primarily of humans (anthropophilia) and (vi) susceptibility to the Plasmodia that affect humans. In addition, longevity was also a crucial criterion for transmission (Dalla Bona & Navarro-Silva 2010). Overall, the lifetime of a primary vector is longer than that of a secondary one, increasing its probability of becoming infected and infective. In addition, the transmission capacity of a vector has been shown to be affected by its immunity response level against the sporogonic cycle of Plasmodium (Arévalo-Herrera et al. 2011). In the Americas, few species of Anopheles fulfil all the characteristics of a primary vector. The Anopheles (Nyssorhynchus) darlingi Root, 1926 stands out due to its marked anthropophilia and endophilia, followed by Anopheles (Nyssorhynchus) nuneztovari Gabaldón, 1940 and Anopheles (Nyssorhynchus) albimanus Wiedemann, 1920. However, using a novel approach (i.e., that of Sinka et al. 2010) that characterises the dominant vector species based on several bionomic aspects that are useful to direct control measures, it is plausible to include species such as Anopheles (Anopheles) pseudopunctipennis Theobald s.l., Anopheles (Anopheles) quadrimaculatus Say, Anopheles (Anopheles) freeborni Aitken, Anopheles (Nyssorhynchus) aquasalis Curry, Anopheles (Nyssorhynchus) marajoara Galvão & Damasceno and Anopheles (Nyssorhynchus) albitarsis Lynch-Arribálzaga s.l. as primary vectors. Primary malaria vector species - Colombia has a high number mosquito species compared to neighbouring countries and is a potential hotspot for malaria endemicity (Foley et al. 2008). According to González and Carrejo (2007), five subgenera of Anopheles, with between 40-47 species, are found in Colombia (Table I). Of these, only three Nysorrhynchus species (An. darling, An. albimanus and An. nuneztovari) are primary vectors of malaria (Table II). However, in light of the criteria proposed by Sinka et al. (2010), species such as An. (An.) pseudopunctipennis and Anopheles (Kerteszia) neivai (Howard, Dyar & Knab, 1917)display ecological and epidemiological characteristics in Colombia (Table II) that concur with the definition of a primary rather than a secondary vector. An. (Nys.) darlingi - This species, an inhabitant of jungle and forest environments, is mainly distributed in regions less than 500 m above sea level (masl) in areas with high relative humidity, precipitation of more than 1,000 mm and temperatures between 21-28ºC (Rubio-Palis & Zimmerman 1997). Geographically, its population extends along the length and width of the Andes, from Argentina-Colombia, where it is also found to the west of this mountain range. It reappears in Panamá (Loaiza et al. 2009) and extends to the north in Honduras, El Salvador, Guatemala and Belize, reaching southern Mexico (Manguin et al. 1996, 1999). There is considerable biological variation from country to country (Quiñones & Suarez 1990, Manguin et al. 1999). For example, in northern Colombia, An. (Nys) darlingi is found between the foothills and the coastal zone in areas with over 4,000 mm yearly precipitation (Suarez et al. 1990) (Fig. 1), while in Venezuela it is found in areas with lower precipitation (1,276 mm) in the south, near the border with Brazil and Guyana (Moreno et al. 2007). It is unable to survive in dry or arid areas (Sinka et al. 2010). In Venezuela, the abundance peaks for An. (Nys.) darlingi populations and the transmission of malaria occurred after maximum precipitation (Magris et al. 2007). In contrast, transmission occurred during dry periods in Brazil (Camargo et al. 1996, Galardo et al. 2009). In Peru, their greatest abundance is associated with deforested sites and is related to the availability of breeding sites. Annual infectious bites increase in proportion to increases in deforestation and become reduced when there is maximum precipitation and when the temperature drops (Vittor et al. 2006). Throughout its range, An. (Nys.) darlingi shows great morphological, chromosomic, isoenzymatic and genetic variability to the point that it may be considered part of a species complex (Manguin et al. 1995, Charlwood 1996). At least six major population groups are recognised (Pedro & Sallum 2009). However, recent analysis of the entire mitochondrial genome cytochrome c oxidase subunit I (COI) did not support a speciation process in An. (Nys.) darlingi (Moreno et al. 2010). For instance, in Colombia, microsatellite and COI analyses showed low and moderate genetic differentiation, respectively (Gutiérrez et al. 2010b), and supported panmictic populations (González et al. 2007) in this species. In natural environments, An. (Nys.) darlingi occupies diverse habitats but is almost always limited to small, unpolluted bodies of fresh water, such as streams, stream banks and lagoons located in shady areas containing floating aquatic plants (e.g., Pistia stratiotes) (Olano et al. 2001, Brochero et al. 2005, Rubio-Palis et al. 2005) or floating mats of wood pieces, dead leaves and flower and seed debris (Manguin et al. 1996). In disturbed areas, cisterns, ditches, gutters, artificial wells and aquaculture ponds are exploited as breeding sites, independent of whether these sites are partially or totally shaded, contain a substantial level of litter or are poor in vegetation and nutrients (Tadei et al. 1998), but requiring a pH of between 6.0-8.0. In the Peruvian Amazon, An. (Nys.) darlingi larvae were found in breeding sites located in pastures associated with shrub or secondary forest (Vittor et al. 2006). Although it shares its ecological niche with other anophelines, it is generally the predominant species where ever it is found (Rubio-Palis & Zimmerman 1997). Although An. (Nys.) darlingi is associated with settlements near jungles and forest environments, it has been found in more semi-urban environments. This is the case in the peri-urban area of the city of Quibdó (Chocó, Colombia), a site of urban malaria infections, and An. (Nys.) darlingi, together with An. (Nys.) nuneztovari, is considered to be involved in malarial transmission (Ochoa & Osorio 2006). Another important aspect regarding An. (Nys.) darlingi is the plasticity of its pattern of biting activity. This varies according to its geographical location. For example, in the Amazonian areas of Surinam, Colombia, Venezuela, Peru and Brazil, biting typically reaches a single peak before midnight (Elliot 1972, Hudson 1984, Rozendaal 1989, Quiñones et al. 1992, Vittor et al. 2006, Magris et al. 2007). However, in other regions of Brazil (Charlwood & Wilkes 1979, Forattini 1987, Roberts et al. 1987, Lourenço-de-Oliveira et al. 1989, Tadei et al. 1998) and Colombia (Rodríguez et al. 2009), biting activity displayed a bimodal peak: one peak at the beginning of the night and other peak at dawn. In contrast, in French Guyana (Pajot et al. 1977) and in other places in Brazil (Freitas-Sibajev et al. 1995), biting activity reached three peaks (sunset, midnight and dawn). The other essential issue to mention is the variable biting habits of An. (Nys.) darlingi. A classic study by Rachou (1958) (quoted by Zimmerman et al. 2006) demonstrated that the human blood index (HBI) for this species ranged between 0.120-0.853 in the state of Pará (Brazil). The HBI for Bolivian and Colombian An. (Nys.) darlingi was within this range (WHO 1960). Based on these observations, Linthicum (1988) hypothesised that one of the major characteristics of An. (Nys.) darlingi is its marked anthropophilia, an aspect verified by Oliveira-Ferreira et al. (1992) and Zimmerman (1992). However, in a comparative study, Zimmerman et al. (2006) found that anthropophilic habits might vary according to host availability and the site of capture. Indeed, they found that An. (Nys.) darlingi appeared to be highly anthropophilic, with variable rates between 0.131-0.405 when the captures were made from humans in three riverine Brazilian villages of state of Amapá (AP). Surprisingly, enzyme-linked immunosorbent assay (ELISA) analysis revealed a higher rate of bovine positive (56.9%) than human positive blood (11.7%). Oliveira-Ferreira et al. (1992) recorded this species as a mosquito capable of feeding on both humans and cattle. An. (Nys.) darlingi has been implicated as a malaria vector in various South American localities (Mirabello et al. 2008). Its infection rates range between 0.5-2.5% (Oliveira-Ferreira et al. 1990, Tadei & Dutary-Thatcher 2000, Magris et al. 2007). In the Amazon Region, just as in the Venezuelan Orinoco, its abundance has correlated positively with malaria cases (Magris et al. 2007). Recently, in Peru, the vector was observed naturally infected with Plasmodium vivax and Plasmodium falciparum (Flores-Mendoza et al. 2004) and in Venezuela with Plasmodium malariae (Magris et al. 2007), whereas in AP it was found harbouring the three human species of malaria (Galardo et al. 2007). In Colombia, An. (Nys.) darlingi is the major vector of P. falciparum in the humid tropical forest of Chocó, in the western foothills of the Andes (Suarez et al. 1990, Rubio-Palis & Zimmerman 1997) and in the Eastern Plains (Brochero et al. 2005). In Puerto Leguizamo, state of Putumayo, it is present and considered to be the local vector of P. falciparum (Quiñones et al. 2006). More recently, in department of Córdoba, An. (Nys.) darlingi was observed to be naturally infected with P. vivax (Gutiérrez et al. 2009). An. (Nys.) darlingi is also a vector in Quibdó even though it has a low abundance (Ochoa & Osorio 2006). In summary, An. (Nys.) darlingi is, without a doubt, the most efficient malaria vector in Colombia (Brochero et al. 2005, Quiñones et al. 2006), as it is in the rest of the Neotropics (Foote & Cook 1959, Manguin et al. 1999, Galardo et al. 2007), with exception of a region of the Brazilian Amazon, where its role as the primary vector may be replaced by An. (Nys.) marajoara as consequence of changes in land use (Conn et al. 2002). This efficiency is due to its high degree of anthropophilia, its diverse habits of endophilia and endophagy, its adaptability to domestic environments and its dispersal, in combination with its high degree of natural (1%) and experimental (23%) susceptibility to infection (Flores-Mendoza et al. 2004, Bharti et al. 2006) with human Plasmodia species. An. (Nys.) albimanus - This is one of the most widely studied anophelines because it is one of the major vectors of human Plasmodia in Central America, northern South America and the Caribbean. In addition, it is the most well represented of Nyssorhynchus species in laboratory colonies. Frederickson (1993) provides an excellent review of its geographic distribution, ecological characteristics and epidemiological importance. This species has been found primarily in low coastal lands less than 400 masl and has a marked preference for areas below and up to 100 masl (Frederickson 1993), although it has been reported at higher elevations (Guatemala at 1,372 and 1,200 masl, Mexico at 1,941 masl and Colombia at 1,000 masl) (Vargas & Martinez-Palacios 1955, Frederickson 1993, González & Martínez 2006). It is found from Texas-Venezuela and on most of the Caribbean islands (Faran 1980). Its absence is notable in Trinidad and the Lesser Antilles (Rubio-Palis 2000) as well as in Brazil. On the Pacific coast, it is found from Mexico (Rubio-Palis & Zimmerman 1997) to northern Peru (Calderón et al. 1995). Its geographic distribution coincides with areas that have two rainy seasons with precipitation greater than 1,000 mm (although there is an exception in Guatemala) (Ogata et al. 1992), high relative humidity and a monthly variation in temperature of between 22-29ºC (Ogata et al. 1992, Rubio-Palis & Zimmerman 1997, Loaiza et al. 2010). At the national level, with few exceptions, this Anopheles species is predominant in localities along the Pacific coast (Olano et al. 2001) and, to a lesser degree, along the Caribbean coast, extending to the Isla Fuerte (Bejarano 2003) and the Archipelago of San Andrés (Olano et al. 2001). Its distribution (Fig. 2) and abundance seem to be determined by the availability of breeding sites. Along the Naya River (Valle del Cauca), it is primarily found along the coastline but completely disappears at 100-200 masl, likely due to the absence of breeding sites (Solarte et al. 1994). Nevertheless, it has colonised some areas with higher altitudes, such as those reported in Cauca (González & Martínez 2006) and Valle (R González, unpublished observations), around 1,000 masl. In spite of the apparent plasticity of An. (Nys.) albimanus populations, they are cytologically and morphologically homogeneous (Keppler et al. 1973, Faran 1980, Frederickson 1993, Molina-Cruz et al. 2004) with genetic flow between populations. However, morphological variation in An. (Nys.) albimanus larvae has been observed in Ecuador (R Wilkerson, unpublished observations). Recently, evidence from DNA analyses using nuclear and mitochondrial markers support a single species of An. (Nys.) albimanus throughout its geographical range (Loaiza et al. 2010) in Central and South America. However, in Colombia, two distinct lineages were observed in the Caribbean and Pacific regions using COI sequences (Gutiérrez et al. 2009). An. (Nys.) albimanus breeding sites are generally exposed to the sun, but larvae can tolerate a certain degree of shade (Faran 1980, Frederickson 1993) and salinity (up to 37% sea water) (Frederickson 1993). The larval populations commonly increase with the rains and decrease during dry periods, during which they survive in mangroves, lagoons and coastal canals (Frederickson 1993). In Colombia, this species has been found in a great variety of aquatic (lakes, streams, wells) and semi-aquatic (gutters, animal and vehicle tracks, cavities in tree trunks, rocks or animals in mangroves) habitats (Marten et al. 1996). Often, its presence has been directly associated with the presence of hydrophytes, such as Pistia sp., Naiais sp., Chara sp. and Utricularia sp. (Torres-Estrada et al. 2005), and with Dalbergia, Costus, Cyperus, Wedelia, Desmodium, Orizam, Abutilon, Calathea and Utricularia (Solarte et al. 1994, Torres-Estrada et al. 2005). It has likewise been observed in urban areas (Olano et al. 1997) and areas disturbed by human activity. In various south-western Colombia localities, its primary breeding sites consisted of mining excavations, aquaculture pools and lagoons (Quiñones et al. 1987, Olano et al. 1997) as well as excavations left by brick pits (González & Martínez 2006). In some areas along the Colombian Pacific coast, the anthropophilic activity of An. (Nys.) albimanus was variable, with at least two peaks: one towards midnight and a second one of less intensity before dawn in both intra and peri-domicile areas (Olah & Montoya 1985, Quiñones et al. 1987, Frederickson 1993, Solarte et al. 1996, Rubio-Palis 2000). Solarte et al. (1996) established that on the Naya River nearly 19% of mosquitoes entered houses to feed, a behaviour later confirmed by Olano et al. (1997) for the urban area of Buenaventura. In this area, the peak of greatest haematophagic activity is located outside of houses between 6:00-9:00 pm. Then, at midnight, there is a second peak inside of houses. Finally, between 05:00-06:00 am, this activity increased again in the peri-domicile areas (Olano et al. 1997). However, in Peru, the period of maximum activity was found to be between 8:00-10:00 pm (Calderón et al. 1995), extending to 11:00 pm in Mexico and Nicaragua (Frederickson 1993). The adult abundance of An. (Nys.) albimanus was, with few exceptions, directly related to high precipitation (Frederickson 1993) and nearly disappeared during the dry season (Dieguez et al. 2002). However, in the urban areas of Buenaventura, where it is considered the most important vector, the increase in bite rate was related to the reduction in rainfall (Olano et al. 1997). There was a concurrent decrease in malaria cases simultaneous to a reduction in bite rate in Buenaventura. This finding is in contrast with the malaria outbreak in La Guajira (Porras et al. 2001), where a direct relationship was found between a reduction in rainfall and an increase in cases of malaria. Recent studies have related the El Niño-Southern Oscillation Event, which affects global climatic conditions every two-seven years, to the increase in malaria cases transmitted by An. (Nys.) albimanus along the Colombian Pacific coast (Poveda et al. 2001). Adults of An. (Nys.) albimanus have a short flight range and are endophagic and exophagic, the latter being the prevalent characteristic (Solarte et al. 1996, Olano et al. 1997). In El Salvador, mark, release and recapture studies demonstrated that adults travel 3 km in 6.5 days, although (Curry 1925) a greater dispersal range has been estimated in Panama (19 and 32 km), possibly aided by wind. Olano et al. (1997) established that in Buenaventura, this species exhibits low exophilia in corrals and stables in an area where the availability of cattle was scarce; instead it displayed anthropophilic habits. This finding contrasted with literature reporting that donkeys, horses, cows and dogs are highly preferred (Frederickson 1993). The biting activity of An. (Nys.) albimanus between 7:00-11:00 pm (primarily outdoors) is possibly of greatest epidemiological significance in the area because many people are outside and are exposed to the bites during this period interval. In field samples collected from Central American countries, the proportion of gravid females obtained varies between 37.8-70%. It is thought that gravid females, prone to transmit infection, are linked to this range (OPS 1996). An. (Nys.) albimanus has been observed to be naturally infected with both P. falciparum (Herrera et al. 1987, Mekuria et al. 1991, Arredondo-Jiménez et al. 1995) and with P. vivax (Calderón et al. 1995, Gutiérrez et al. 2008), although its populations vary considerably with regards to their vectorial competence, biting habits and feeding preferences throughout the species' geographical range. It is the primary vector of malaria in southern Mexico (Arredondo-Jiménez et al. 1995) and in various countries of Central and South America (Faran 1980, Frederickson 1993, Rubio-Palis & Zimmerman 1997), including Colombia, where it is the principal vector in rural and peri-urban areas of the Pacific coast and, possibly, on the Caribbean coast (Gutiérrez et al. 2008). An. (Nys.) nuneztovari - An. (Nys.) nuneztovari is widely distributed in eastern Panama and northern South America, primarily in the eastern foothills of the Andean Mountains, in the Amazon and in the grasslands of the Colombo-Venezuelan plains (Faran 1980, Rubio-Palis & Zimmerman 1997, Sierra et al. 2004) (Fig. 3). In Colombia, it was found that aquaculture ponds are one of this species' most frequent breeding sites (López & González 1994). For example, in Cimitarra (Department of Santander), ponds represented approximately 81% of the breeding sites that, in addition to being permanent, were peri-domicilary, completely exposed to the sun and contained emerging vegetation, particularly grasses (Brochero et al. 2006). López and González (1994) found that the phytoplankton and the water-vegetation interphase of aquaculture ponds were fundamental to the survival of the larvae of this species. It has also been observed that deforestation produced by monoculture favours the presence of these breeding sites (Brochero et al. 2006). Lagoons and rainwater pools are sites that harbour An. (Nys.) nuneztovari larvae in Quibdó and Buenaventura (Valle del Cauca) (Olano et al. 2001, Sierra et al. 2004, Ochoa & Osorio 2006). An. (Nys.) nuneztovari s.l. exhibits variation in its behaviour patterns: in Suriname it is an exophilic species and bites human at sunset (Panday 1977); in Brazil, it is more zoophilic and exophagic, with a stronger association with sunset and peri-domicile habits than in Venezuela, where it exhibits marked anthropophilia, endophagia and endophilia (Gabaldón 1981, Olano et al. 1997, Brochero et al. 2006). In the Rio Ene Valley, a hyperendemic malarious region of Peru, it was found to be active from 11:00 pm-04:00 am, with a peak between 03:00-04:00 am (Hayes et al. 1987). In a pioneering study in northern Colombia, Fleming (1986) observed populations of this species that exhibited intra-domicile patterns with high anthrophilia and an activity peak late at night. In recent studies, Parra-Henao and Alarcón-Pineda (2008) recorded an exophagic behaviour in Córdoba, in contrast with Brochero et al. (2006) in Cimitarra, who observed an endophagic pattern. The different behaviour patterns determined for this species have been related to the three cytotypes. Type A [Anopheles (Nyssorhynchus) goelddi Rozeboom & Gabaldón], present in Brazil, is zoophilic and exophagic, while Types B and C, which are both found in Colombia and Venezuela, are predominately both anthropophilic and endophagic (Scarpassa & Tadei 2000). Endophagic-exophilic differences over the species' geographic range influence the use of residual insecticides to control malaria. In Venezuela, it was found that the adult abundance of An. (Nys.) nuneztovari increased two months after rainfall peaks and was additionally related to the rain accumulated during that period (Scorza et al. 1981). In Bajo Calima, Colombia, the adults displayed a similar bimodal pattern, with a high peak between June-July and secondary one between January-February (Fajardo et al. 1983). An. (Nys.) nuneztovari has long been recognised as a major vector in Colombia, Peru and Venezuela (Elliot 1972, Hayes et al. 1987, Rubio-Palis et al. 1992). It has been observed to be naturally infected with P. vivax (Rubio-Palis et al. 1992) and P. falciparum (de Arruda et al. 1986). It is one of the species that transmits Plasmodium in Bajo Calima and Citronella (Buenaventura) (Rey & Rengifo 1950, Fajardo et al. 1983, Fajardo & Alzate 1987), areas where it is dominant both in adult and immature forms (Olano et al. 1997). It is likewise a vector in Tierralta (Córdoba) (Gutiérrez et al. 2009) and Tibú (Norte de Santander). Furthermore, due to its marked anthropophilia and endophilia, as well as its abundance and vectorial background in other places, it is suspected of being a vector together with An. (An.) pseudopunctipennis s.l. and An. (Nys.) darlingi in Cimitarra (Posso et al. 2003, Brochero et al. 2006). Elliot (1972) proposed that these populations were allopatric after considering differences in host preferences and biting patterns among An. (Nys.) nuneztovari populations in Colombia and Venezuela compared to those in the Brazilian Amazonia. This proposal was confirmed by several morphological, chromosomal, molecular, behavioural and genetic flow studies (Fritz et al. 1994, Hribar 1994, Linley et al. 1996, Conn et al. 1998, Tadei et al. 1998, Scarpassa et al. 1999, Scarpassa & Tadei 2000). Most recently, it was estimated that An. (Nys.) nuneztovari represents a complex of at least five lineages (Mirabello & Conn 2008), as was suggested some years ago (Conn 1990, Conn et al. 1993, Sierra et al. 2004, Brochero et al. 2006). Often, there is a misidentification of An. (Nys.) nuneztovari, Anopheles (Nyssorhynchus) dunhami and An. (Nys.) goelddi because of the absence of high-quality morphological keys. At present, the An. (Nys.) nuneztovari complex is comprised of the following: An. (Nys.) goeldii, (An. (Nys.) nuneztovari A and An. (Nys.) nuneztovari C (Sierra et al. 2004). According to Sierra et al. (2004), An. (Nys.) nuneztovari populations located along the eastern margin of the Andean Mountains in Norte de Santander and the western margin in Chocó and Valle del Cauca correspond to a unique genetic species. This is despite differences in its karyotype that have been reported by Conn et al. (1993). Recently, Ruiz et al. (2010) reported the presence of the related species An. (Nys.) dunhami in the Amazon and Calado et al. (2008) were able to identify An. (Nys.) goeldii in AP based on similarity to An. (Nys.) nuneztovari. Both species were previously identified as An. (Nys.) nuneztovari using female morphology. In other Colombian regions, An. (Nys.) nuneztovari has been misidentified as several Anopheles species (Ramos et al. 2008), namely Anopheles (Nyssorhynchus) oswaldoi Peryassú s.l., An. (Nys.) nuneztovari, An. (Nys.) aquasalis, Anopheles (Nyssorhynchus) benarrochi Gabaldón, Cova-García & López s.l., Anopheles (Nyssorhynchus) evasae, Anopheles (Nyssorhynchus) strodei and Anopheles (Nyssorhynchus) trinkae. The bionomic variation in An. (Nys.) nuneztovari reported in South America requires clarification to establish the true distribution of the species. Secondary malaria vector species - Seven other species of anophelines, An. (An.) pseudopunctipennis, Anopheles (Anopheles) punctimacula Dyar & Knab s.l., Anopheles (Anopheles ) calderoni (Wilkerson 1991) Anopheles (Anopheles) neomaculipalpus (Curry 1933), Anopheles (Kerteszia) lepidotus (Zavortink 1973), An. (Ker.) neivai, Anopheles (Nyssorhynchus) rangeli Gabaldón, Cova-García & Lópes and An. (Nys.) oswaldoi (Carvajal et al. 1989, Quiñones et al. 2006) act as secondary vectors, are suspected of being associated with transmission or have been identified as having regional or local importance (Olano et al. 2001, Moreno et al. 2005, Quiñones et al. 2006, Gutiérrez et al. 2008). With the exception of An. (Nys.) rangeli, all of the above species are now recognised to be members of species complexes, making it difficult to associate previous information regarding their bionomic or vectorial status with a single species within each species complex. The list of species (and vectors) is constantly being reconstructed and updated as the genus Anopheles has several sibling species complexes (Krzywinski & Besansky 2003), some of which are found in Colombia, e.g., An. (Nys.) benarrochi (Quiñones et al. 2001, Ruiz et al. 2005), An. (Nys.) marajoara s.l. (Brochero & Quiñones 2008) and An. (Nys.) oswaldoi (Ruiz et al. 2005). An. (An.) pseudopunctipennis - Biochemical and molecular analyses and crosses established that this species is a complex composed of at least three "species": one from southern USA, Mexico and Guatemala, another from Central America, Belize and South America and the third from Grenada Island (Dantur et al. 2010). However, Manguin et al. (1995) argued that there is a single species with three widely distributed populations: one in the United States, Mexico, Guatemala, and Belize, another in Colombia, Ecuador, Peru, Chile and Argentina and, finally, a locality type present on Grenada Island. The neotype was described by Rueda et al. (2004). To fix the nominotypical name, a re-description was performed that modified both the type locality and the neotype. An. pseudopunctipenis has been observed to occupy forest and anthropic environments and to prefer transition areas and regions close to jungles in Salta (Argentina) (Dantur et al. 2005). In Peru, it is highly anthropophilic in both the intra and peri-domicile environments, with activity peaking between 6:00-12:00 pm and 04:00-06:00 am (Calderón et al. 1995), while in Venezuela it is active both day and night and exhibited a facultative anthropophilic behaviour (Elliot 1972, Fleming 1986, Olano et al. 2001). In Venezuela, An. (An.) pseudopunctipennis prefers to bite animals instead of humans (Berti et al. 1998). In Mexico, larvae of An. (An.) pseudopunctipennis s.l. are abundant along river banks during the dry season (Fernandez-Salas et al. 1994). In contrast, during the rainy season, it appears that the river carries them away and eliminates them (Savage et al. 1990, Rejmánkova et al. 1991); a correlation was found between the number of larvae and the rainfall of the previous month. Larvae were positively associated with green (Spirogyra) and filamentous algae (Savage et al. 1990, Rejmánkova et al. 1991) along river banks, specifically where there were very slow currents or in small pools resulting from river overflow (Berti et al. 1998). An. (An.) pseudopunctipennis s.l. has been observed to be naturally infected with P. vivax (Hayes et al. 1987). Infection with P. vivax has been established by ELISA in the Valle del Bajo Chira, in the localities of Salitral and Querecotillo in Peru (Calderón et al. 1995), and with P. falciparum in Panama (Loaiza et al. 2008). Additionally, it is a suspected vector for P. falciparum in areas where the coast abruptly transforms into mountains as occurs in the western foothills of the Andean Mountains in Peru, in Chiapas, Mexico (Hoffman 1932) and in foothills of lesser altitude (< 1,000 m) from Colombia-Argentina (Rubio-Palis & Zimmerman 1997). In these areas at altitudes over 600 m, it is frequently the only vector present (Manguin et al. 1995). This is likewise the situation in areas near Cali (Valle del Cauca), where An. (An.) pseudopunctipennis s.l. has been incidentally associated with epidemic outbreaks (Ocampo et al. 2000). An. (An.) punctimacula s.l. - This species has been considered to be a secondary vector in the coastal regions of Central America and northern Colombia (Rubio-Palis & Zimmerman 1997). Studies in Colombia and Ecuador have shown that most reported incidences of An. (An.) punctimacula were in fact An. (An.) calderoni, a member of the An. (An.) punctimacula s.l. group. Anopheles (Anopheles) malefactor Dyar & Knab (Wilkerson 1990) and Anopheles (Anopheles) shanonni Davis (Wilkerson et al. 1997, González et al. 2010) are the two other species in this group and they may be present in Colombia. The role of each of these species in malarial transmission is unknown. An. (An.) calderoni has been identified in several malarious eco-regions (coast, foothills and high valleys) of South America, including areas of Ecuador, Peru, Venezuela and Colombia (González et al. 2010). In Peru, it was found from coastal areas to 1,080 masl; in some instances, it competed in abundance with An. (Nys.) albimanus and An. (An.) pseudopunctipennis. An. (An.) calderoni has been incriminated as a secondary vector of P. vivax (Calderón et al. 1995). An. (An.) calderoni is a valid species (González et al. 2010) independent from An. (An.) punctimacula and An. (An.) malefactor (Wilkerson 1991). It is likely that a large portion of the data for An. (An.) puntimacula from Colombia (Olano et al. 2001), Ecuador (Levi-Castillo 1949) and Peru actually corresponds to An. (An.) calderoni. In Peru, the reproductive cycle of this species is associated with irrigation water channels and fish farms in crop culture in the Peruvian Amazon (Vittor et al. 2006). In Colombia, An. (An.) calderoni exhibits a nocturnal, anthropophagic behaviour. Both peri and extra-domiciliar catches produced a unimodal peak in the first hours of the night, although this peak was variable in other localities (7:00-8:00 pm and 10:00-11:00 pm) (Lucumí et al. 2009). An. (An.) neomaculipalpus - This species is widely distributed from Mexico-Argentina (Knight & Stone 1977), but little is known about its bionomy and ecology due to its scarcity. In Venezuela, it has been reported to have a high anthropophilic index (50%) in areas where An. nuneztovari was the major vector (Rubio-Palis et al. 1994), and in Cimitarra it was recorded to be attracted to a Shannon trap in low density in the extra-domicile environment (Brochero et al. 2006). An. (An.) neomaculipalpus from Venezuela was observed to be infected with P. vivax with a sporozoitic rate greater than those recorded for An. (Nys.) marajoara (0.84 vs. 0.82) or for An. (Nys.) darlingi (0.27), the major vector in the area (Moreno et al. 2005). In Colombia, it was observed to be naturally infected with P. falciparum (Herrera et al. 1987). Similarly, Gutiérrez et al. (2009) recorded two specimens positive for infection with P. falciparum in Turbo. An. (Ker.) lepidotus - In Colombia, this species was confused with Anopheles (Kerteszia) boliviensis by Bates (1943), who recorded that, in the Department of Meta, it was a very common mosquito with morning and evening activity peaks and that it was limited to tree canopies. Bates (1943, 1944) found that this species had diverse biting habits, observing that it attacked humans during the day, but horses at night. Later findings revealed that this species is very common in humid, pre-mountain forest areas in Tolima that are characterised by abundant epiphytes, including the bromeliad Vriesea capituligera, which serves as a breeding site (Quiñones et al. 1984). According to these authors, the species had daytime habits with population peaks between January-June and a maximum haematophagy and marked exophagy between 3:00-6:00 pm. Based on epidemiological evidence, the authors affirmed that An. (Ker.) lepidotus fulfils several of the requirements for consideration as an important vector in the transmission of P. vivax in Tolima. However, the species name has recently been called into question and it is the subject of on-going morphological and molecular studies (Escobar et al. 2010, Bruce Harrison, RC Wilkerson, unpublished observations). An. (Ker.) neivai - This species is primarily found associated with epiphytic and terrestrial bromeliads: Vriesea (Vriesea gladioliflora, Vriesea sanguinolenta) and Guzmania (Guzmania sanguinea, Guzmania sherzeriana var. sherzeriana and Guzmania musaica) (Murillo et al. 1988a, Solarte et al. 1994), although it was additionally found in tree holes. In Panama, Trapido and Galindo (Zavortink 1973) observed this species to occupy arboreal areas of the rain forest and ground level in deciduous forests. Two studies carried out on the Colombian Pacific coast demonstrated that it was abundant in disturbed forests predominantly populated by mangroves (Rhizophora mangle, Avicenia nitida and Mora megistosperma), where there was a characteristic increase in bromeliads (Astaiza et al. 1988, Solarte et al. 1994). Solarte et al. (1994) established its presence in transitional areas between mangroves and forests where Euterpe cuatrecasana, Pterocarpus officinalis, Symphonia globulifera and M. megistosperma predominated and where the greatest number of bromeliads (1.1-2.2 per m2) were found. It is interesting to note that 87% of the larvae were found within 0-5 m of the ground and the greatest larvae index was found within 0-2 m (Solarte et al. 1994). In these areas, which are characterised by the absence of stables or domestic animals, the species is typically active over the entire day (Solarte et al. 1996); however, at dusk the population becomes dispersed towards nearby towns where humans are present. Because of its marked exophilia and the lack of adequate resting and oviposition sites in the towns, most individuals later return to the mangroves, where breeding sites were abundant (Astaiza et al. 1988). An. (Ker.) neivai was thought to be anthropophilic, but its feeding preferences have not been unequivocally demonstrated. Nevertheless, in Charambirá, Colombia An. (Ker.) neivai was found to be attracted to humans and biting activity peaks at twilight from 05:30-06:30 am and 6:00-7:00 pm. Additionally, there is a low level of intra-domicile activity that extends all night. In Charambirá, it was established that An. (Ker.) neivai activity began at 10 lux units (i.e., equivalent to 1 lumen/m²), achieved a maximum at around three units and ended when luminosity descended to 0.1 lux unit (Murillo et al. 1988b). The first peak was dominated by long-lived females, generally multiparous (i.e., more than 3 times), with a greater opportunity to transmit Plasmodium (Astaiza et al. 1988). This observation was validated by Murillo et al. (1988b), who additionally analysed the age-range structure of this species based on the gonadotropic state of females captured in the field. These authors postulated that the females had a long life span based on the multiple gonadotropic cycle observed. An. (Ker.) neivai did not present intra-domicile activity but exhibited an extra-domicile activity peak between 6:00-8:00 pm in other locations on the Colombian Pacific coast such as in Zacarias (a town near Buenaventura) (González et al. 1997) and marked daytime activity within forests near mangroves where its breeding sites predominate such as in the Naya River area (Solarte et al. 1996). Several epidemiological field studies point to An. (Ker.) neivai as a potentially important vector of human malaria at various sites between Buenaventura-Chocó along the Colombian Pacific coast (Lee & Sanmartin 1967, Muñoz & Parra 1984, Olah & Montoya 1985, Hurtado & Solarte 1986, Astaiza et al. 1988). This possibility has been strengthened by the observation of mosquitoes naturally infected with both P. vivax and P. falciparum simultaneously (Herrera et al. 1987, Carvajal et al. 1989, Gutiérrez et al. 2008) as well as by the results of experimental infections with Plasmodia (Carvajal et al. 1989) and the results of sociological studies (Sevilla-Casas 1993). An. (Nys.) rangeli - This species is distributed east of the Andes from Colombia to Bolivia, including Venezuela and Brazil (Hayes et al. 1987, Póvoa et al. 2001, Fritz et al. 2004, Brochero et al. 2005). In Colombia, larvae have been found in fish culture ponds, streams, morichales (mirity palms), lakes and brooks (Brochero et al. 2005). Biting activity occurred around houses from 6:00-7:00 pm (Brochero et al. 2005). Although it was found in Peru to be naturally infected with unspecified Plasmodium species (0.4%) (Hayes et al. 1987) and to be infected with P. malariae in Brazil (Póvoa et al. 2001), its vectorial status in Colombia has been questioned. While Brochero et al. (2005) did not consider it to be an important vector, Quiñones et al. (2006) observed this species to be naturally infected with P. vivax (8.7%) in southern Colombia and concluded that this species was involved in malaria transmission in this region. An. rangeli shows, in general, discriminant morphological characteristics in both its immature stages and in the adult female. However, in some cases, and particularly when this species is found sympatric with An. nuneztovari, their correct determination using only adult female morphology can be difficult. In the region of San Pedro de Urabá (Antioquia, Colombia) (Zapata et al. 2007), mosquitoes were identified from morphological characteristics as "possibly" An. oswaldoi, An. rangeli and An. strodei, but internal transcribed spacer-2 (ITS-2) restriction fragment length polymorphism patterns from type specimens confirmed that all were An. nuneztovari. Due to the high interspecific similarity between An. rangeli and An. nuneztovari, the use of a molecular tool is necessary for the correct taxonomic determination of these species. An. (Nys.) oswaldoi s.l. - This is a well-recognised species complex (Marrelli et al. 1999). These authors reported four species from Brazil and Peru based on data from the ITS-2 sequences. In Colombia, An. (Nys.) oswaldoi is a complex of at least two species (F Ruiz, unpublished observations). One of these, i.e., An. (Nys.) oswaldoi B, was observed to be naturally infected with P. vivax in southern Colombia, in the state of Putumayo, with an infection rate of 0.27% (Quiñones et al. 2006), and in Brazil with P. falciparum (infection rate 3.41%) (Branquinho et al. 1993). An. (Nys.) oswaldoi s.l. has been recorded as a common species in the Colombian Amazon, where it is the major outdoor biting species, while An. (Nys.) darlingi was the major indoor biting species (Rodríguez et al. 2009). Concluding remarks - In this summary, we have presented a review of the major biological characteristics of the three primary vector species of human malaria in Colombia and concluded that they should be considered of highest priority for epidemiologic surveillance. However, in addition to these three principal vector species, our review also seeks to direct attention to the monitoring of other anopheline species that, although neglected, fulfill many of the ecological and epidemiological parameters required for consideration as primary rather than secondary vectors. We concur with Sinka et al. (2010) that there is a need for further study of the bionomics of both primary and secondary vectors along with study of local conditions affecting the transmission of malaria. The research results must then be effectively incorporated into the new policies for malaria elimination (Moonen et al. 2009). Filling the gap between ecologically based and epidemiologically based information continues to be a necessity. Relatively simple bionomic observations that do not require the use of sophisticated technologies will help fill this gap. Here, the geographical location of Colombia and the many ecological conditions favouring the diversity of anopheline species are described. From an epidemiological point of view, three species of the Nysorhynchus subgenus, An. (Nys.) darlingi, An. (Nys.) nuneztovari and An. (Nys.) albimanus, are considered to be primarily responsible for the transmission of Plasmodia to humans in the country. The remaining species may likewise present the characteristics of abundance, sporozootic infection and biting habits sufficiently compatible with the ability to transmit the parasite to humans; however, uncertainties in their taxonomic categorisation and identification must be resolved before their role as vectors can be established in support of subsequent studies of their bionomy and their responses to control measure activities. Another consideration is that the number of species suspected of transmission could increase and the possibility of new primary vectors should be foreseen in the context of the transformation of the environment by various human activities. It is apparent that the impact of human intervention cannot be underestimated as we learn more about the complexity of malaria transmission. It is clear that there are specific dynamics affecting each area and this should be kept in mind from an administrative point of view when considering whether sufficient, appropriate information is available for the design of measures to control the primary vector species. It is essential that the presence of complexes of sister species remain a key factor for consideration when planning for the possibility of resistance and re-occurrence. These factors may trigger a strong cycle in the use of residual insecticides (Brochero & Quiñones 2008). In fact, the most recent study on resistance conducted in Colombia, covering 12 departments with at least three localities in each, revealed that the two major species already present a certain degree of resistance to some of the insecticides used in public health programs (ins.gov.co/?idcategoria=8508#). Furthermore, due to several socio-economic factors (extension of the agricultural frontier, massive displacement, armed encounters, troop movements etc.), a great portion of the nation's territory is now moving towards a scenario with variable epidemiologic conditions that favour the re-emergence of malaria and other illnesses transmitted by vectors. In addition to these social phenomena, we must add the environmental ones that allow species of other subgenera (e.g., Anopheles, Kerteszia) to begin to acquire importance as regional or local vectors, e.g., An. (An.) pseudopunctipennis, An. (Ker.) neivai s.l., An. (Nys.) marajoara. These species exhibit adaptability and will therefore adapt to the disturbances to their habitats (e.g., wooded areas) and/or can become domesticated in urban areas. It would be interesting to study these dynamics (especially those of species replacement) over time and in light of the changes that are occurring in the different biogeographic zones, with the understanding that, as Brochero and Quiñones (2008) expressed: "…species are complex, dynamic organisms in permanent adaptation…". For these reasons, those in charge of public health must pay more attention to the specific interactions occurring between the various participants in the transmission cycle (parasites, vector and host). These interactions should be reviewed from an ecological and evolutionary perspective. The idea of a quick and effective "magic bullet" must be challenged and the conditions explained based on sound scientific data. The natural environment is an active web of complex interactions that react, sometimes in unexpected ways, to the "treatments" (insecticides and parasiticides) applied to it. Our hope is that the studies described here will shed light on the ecology (e.g., feeding habits, breeding sites, and nictameral cycles) of the species and will be a useful contribution to the clarification of taxonomic, systematic and vectorial identification. As a whole, they will contribute to the design, implementation and establishment of precise and well-founded control measures, the cost-benefit of which can be measured in ecological and socioeconomic terms and, above all, guarantee time-space sustainability. Finally, we hope that this review will, in and of itself, encourage the study of the bionomy of the Anopheles in Colombia. Acknowlegements To the Biology Department of the Universidad del Valle and the Instituto de Inmunología del Valle, for the time assigned to JML, RG and YAS for this review, to Dr R Dillon, for a revision of a preliminary version, and to the anonymous evaluators, who highly enriched the quality of the review with their comments and recommendations. References
Copyright © 2011 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc11161f3.jpg] [oc11161t1.jpg] [oc11161f2.jpg] [oc11161t2.jpg] [oc11161f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}