 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz, Vol. 106, No. 7, November 2011 Molecular detection of Schistosoma japonicum in infected snails and mouse faeces using a real-time PCR assay with FRET hybridisation probes Tongjit ThanchomnangI, II; Pewpan IntapanI, III; Pusadee Sri-AroonIV; Viraphong LulitanondI, V; Penchome JanwanI, III; Oranuch SanpoolI, III; Wanchai MaleewongI, III* IResearch
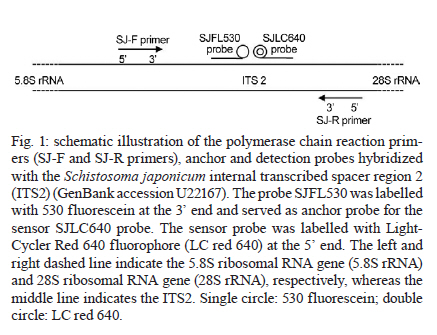
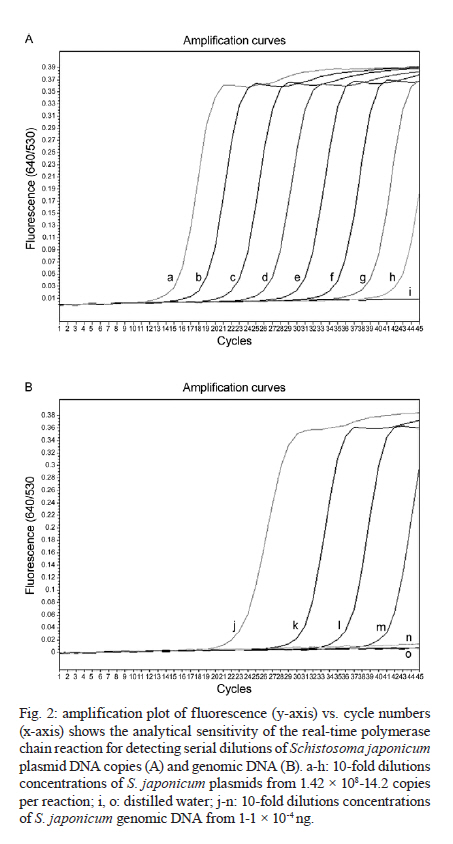
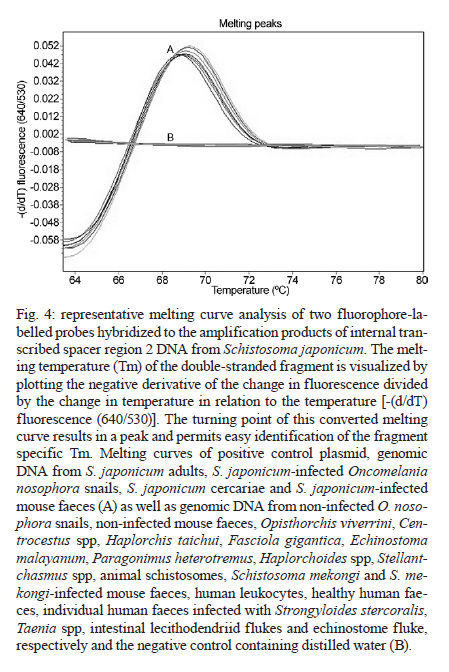
and Diagnostic Center for Emerging Infectious Diseases Code Number: oc11170 ABSTRACT A real-time polymerase chain reaction (PCR) assay with fluorescence resonance energy transfer (FRET) hybridisation probes combined with melting curve analysis was developed to detect Schistosoma japonicum in experimentally infected snails and in faecal samples of infected mice. This procedure is based on melting curve analysis of a hybrid between an amplicon from the S. japonicum internal transcribed spacer region 2 sequence, which is a 192-bp S. japonicum-specific sequence, and fluorophore-labelled specific probes. Real-time FRET PCR could detect as little as a single cercaria artificially introduced into a pool of 10 non-infected snails and a single egg inoculated in 100 mg of non-infected mouse faeces. All S. japonicum-infected snails and all faecal samples from infected mice were positive. Non-infected snails, non-infected mouse faeces and genomic DNA from other parasites were negative. This assay is rapid and has potential for epidemiological S. japonicum surveys in snails, intermediate hosts and faecal samples of final hosts. Key words: Schistosoma japonicum - real-time fluorescence resonance energy transfer PCR - DNA - snails - mice Schistosoma japonicum is an important trematode that causes human schistosomiasis in Asia, including the People's Republic of China, the Philippines and some areas in Indonesia (Zhou et al. 2010). The re-emergence and active transmission of S. japonicum in endemic areas, with a recent infection of approximately 700,000 people, have been reported (Zhou et al. 2007). The S. japonicum life cycle involves a snail intermediate host and a definitive host. Adult schistosomes lay eggs in the mesenteric vein capillaries, which drain the small intestine in the definitive host. The eggs embedded in the intestinal submucosa and mucosa break into the lumen of the bowel and appear in the faeces. In water, the eggs give rise to miracidia, the infective stage of the snail intermediate host. Miracidia penetrate into and migrate through snail tissues to develop into sporocysts, which give rise to cercariae. Humans become infected by coming in contact with water contaminated with cercariae, which actively penetrate into the skin. Parasite transmission requires three factors: surface water contamination with faecal excreta, the presence of specific fresh water snails as the first intermediate host and human water contact (Gryseels et al. 2006). In definitive mammalian hosts, S. japonicum cause feverish syndrome, intestinal diseases and hepatosplenic inflammation in the acute stage of the infection and liver fibrosis in the chronic stage (Gryseels et al. 2006). Human infection has been associated with anaemia (King et al. 2005), hepatosplenomegaly and liver fibrosis, as well as liver and colon cancers (Qiu et al. 2005). Egg detection by stool examination remains the standard method to diagnosis human schistosomiasis japonica. However, it is difficult to differentiate between S. japonicum and Schistosoma mekongi eggs by stool examination; it is not feasible during the prepatent period and it is not sensitive enough to detect mild infections (Doenhoff et al. 2004). Parasite-specific antibody detection by enzyme-linked immunosorbent assay has a high sensitivity and is useful for mass screening, but it has the limitation of a relatively low specificity due to cross-reactivity, and it does not always correlate with an active infection (Doenhoff et al. 2004). The detection of parasite DNA in host-derived material is direct evidence of an actual infection, which is why molecular-based approaches to detect S. japonicum DNA, such as conventional polymerase chain reaction (c-PCR) and loop-mediated isothermal amplification, have been developed (Xia et al. 2009, Kato-Hayashi et al. 2010, Xu et al. 2010). Recently, a SYBR® green-based real-time PCR (Lier et al. 2006, 2008), a minor groove-binding probe-based real-time PCR (Lier et al. 2009), and a TaqMan probe-based real-time PCR (Hung & Remais 2008) were published as sensitive and fast methods that could quantify specific S. japonicum DNA in biological samples and in environmental water. In contrast with previous reports, S. japonicum DNA detection based on the fast, specific and reliable real-time fluorescence resonance energy transfer (FRET) PCR combined with melting curve analysis is promising for detecting an array of parasites from biological specimens (Intapan et al. 2008a, b, 2009a, b). Here, we report the possible applicability of this real-time FRET PCR combined with melting curve analysis to detect S. japonicum larvae in snails and eggs in faecal samples of infected mice. SUBJECTS, MATERIALS AND METHODS Parasite and DNA materials - S. japonicum (Japanese Yamanashi strain) - Experimentally infected Oncomelania nosophora snails and mice, as well as S. japonicum adults and cercariae, were obtained from the Applied Malacology Center, Department of Social and Environmental Medicine, Faculty of Tropical Medicine, Mahidol University, Thailand. All animal experiments were handled according to the Guidelines for Animal Experimentation of the National Research Council of Thailand. S. japonicum (Philippine strain) adults and S. japonicum experimentally infected Oncomelania quadrasi snails, as well as S. mekongi (Loatian strain) adults and S. mekongi infected mice, were also provided obtained from the Faculty of Tropical Medicine, Mahidol University. To evaluate the analytical specificity of the real-time FRET PCR, other parasite DNA was extracted from the adult worms of Opisthorchis viverrini, Haplorchis taichui, Centrocestus spp, Echinostoma malayanum, Fasciola gigantica, Paragonimus heterotremus and Stellantchasmus spp, as well as from metacercariae of Haplorchoides spp, cercariae of animal schistosomes, healthy human faeces, human leukocytes and human faeces infected with Strongyloides stercoralis, Taenia spp, Echinostoma spp and intestinal lecithodendriid flukes. The DNA samples were kept in a DNA bank at -70ºC in the Department of Parasitology, Faculty of Medicine, Khon Kaen University until use. Specimen preparation for real-time FRET PCR - DNA samples were extracted from all non-infected (n = 30) and experimentally infected (n = 30) snails, including their shells, and from snail tissue samples artificially inoculated with S. japonicum cercariae. Each specimen was homogenised with disposable polypropylene pestles (Bellco Glass Inc Vineland, NJ, USA) and extracted using the Nucleospin Tissue kit (Macherey-Nagel GmbH & Co, Duren, Germany). For DNA extraction from mouse faeces, 100 mg of each S. japonicum-infected mouse faeces, non-infected mouse faeces and non-infected mouse faeces artificially inoculated with S. japonicum eggs were thoroughly mixed with 200 µL of normal saline solution (0.85% NaCl in distilled water) and centrifuged at 8,000 g for 5 min. The supernatant was then discarded and the faecal pellet was frozen at -20ºC for 30 min. The frozen pellets were homogenised with disposable polypropylene pestles (Bellco Glass Inc) and extracted using the QIAamp® DNA stool mini kit (Qiagen, Hilden, Germany). The number of S. japonicum eggs in the faeces of infected mice (n = 10) was presented as eggs per gram (EPG) of faeces (ranging from 100-1,100 EPG; geometric mean = 302 EPG). The DNA was eluted in 100 µL of distilled water, 5 µL of which was used for the real-time PCR reaction. Limit of detection - To determine the limit of detection of the real-time FRET PCR, tissues from non-infected snails were separately ground. Next, individual aliquots of one, five and 10 non-infected snail samples were separately inoculated with one, five, 10 and 30 S. japonicum cercariae. Similarly, 100 mg faecal aliquots of non-infected mice were separately inoculated with a serial dilution of one, two, four and eight S. japonicum eggs. These samples were also used for genomic DNA extraction as described above. The resultant DNA samples were then used in the real-time FRET PCR. To evaluate the analytical specificity of the method, genomic DNA from the S. mekongi, O. viverrini, Centrocestus spp, H. taichui, F. gigantica, E. malayanum, P. heterotremus, Haplorchoides spp, Stellantchasmus spp and animal schistosomes, as well as DNA extracted from human leukocytes, negative human faeces and human faeces infected with S. stercoralis, Taenia spp, intestinal lecithodendriid flukes and Echinostoma, were purified as mentioned above. Real-time FRET PCR assay - The LightCycler PCR detection and analysis systems (LightCycler 2.0, Roche Applied Science, Mannheim, Germany) were used for amplification and quantification. The reactions were performed in glass capillaries. The specific primers SJ-F (5'-ATGTTTGATGCGCTCGTA-3') and SJ-R (5'AACTTAGCTTACGGTAGGTG-3') (Proligo, Singapore) were designed to bind to the S. japonicum internal transcribed spacer region 2 (ITS2) sequence (GenBank accession U22167) (Bowles et al. 1995). For amplification detection, the LightCycler FastStart DNA Master HybProbe Kit was used according to the protocol recommended by the manufacturer. Briefly, a pair of adjacent oligo probes were hybridised with the S. japonicum ITS2. One probe was labelled at the 5' end with the LightCycler Red 640 fluorophore (5' Red 640-GTCCGGTCTAGATAGCCAGATTAAGATGCTGCAG-Phosphate 3'; SJLC640 probe) and the other was labelled at the 3' end with a 530 fluorescein (5'CCGTCCGAATAATCCGACCACAGCC - Flou 530 3'; SJFL530 probe) (Tib Molbiol, Berlin, Germany). The probes and primers were designed by LC probe design software (Roche Applied Science). A schematic diagram of the hybridisation analysis of the primers and probes is shown in Fig. 1. When the probes hybridised to the same DNA strand internal to the PCR primers, the probes came in close proximity and produced a FRET (Intapan et al. 2009a). The PCR mixture contained 2 µL LightCycler Faststart DNA Master HybProbe (10 × conc.), 2 mM MgCl2, 0.2 µM SJ-F primer, 0.2 µM SJ-R primer, 0.2 µM SJLC640 probe and 0.2 µM SJFL530 probe. The total reaction volume was 20 µL. The samples were run through 45 cycles of repeated denaturation (10 s at 95ºC), annealing (30 s at 55ºC) and extension (10 s at 72ºC). The temperature transition rate was 20ºC/s. After amplification, a melting curve was produced by heating the product at 20ºC/s-95ºC, which was then cooled to 65ºC, incubated at 65ºC for 3 s and then slowly heated at 0.1ºC/s-80ºC. The fluorescence intensity change was measured throughout the slow heating phase. To evaluate the analytical specificity of the oligonucleotide hybridisation FRET-based technique, DNA extracted from samples other than S. japonicum-infected snail and mouse faecal samples were separately analysed. Each run contained at least one negative control consisting of 5 µL distilled water. For improved visualisation of the melting temperatures (Tm), melting curves were derived as previously described (Thanchomnang et al. 2008). Melting curves were used to determine the specific PCR products, which were confirmed by conventional gel electrophoresis. The cycle number (Cn) was presented as the number of PCR cycles needed for the amplicon fluorescence signal to exceed the detected threshold value. S. japonicum-positive control plasmid - A positive control plasmid was constructed by cloning a PCR product of the S. japonicum ITS2 into the pGEM-T Easy vector (Promega, Madison, WI) according to the protocol of the manufacturer. The PCR products were obtained by c-PCR using the SJ-F and SJ-R primers. The plasmid was propagated in Escherichia coli, and the nucleotide sequence of the inserted gene was sequenced in both directions. The cloned ITS2 nucleotide sequence was identical to the S. japonicum genomic sequence (GenBank accession U22167). Data analysis - The correlation between the worm loads and the Cn was analysed by the Pearson's correlation test. RESULTS Real-time PCR standardisation - The analytical sensitivity of the real-time FRET PCR was evaluated using 5 µL of serial dilutions (1.42 × 10-1.42 × 108 copies) of S. japonicum positive control plasmid in distilled water. The limit of detection of the ITS2 gene target DNA sequence was 1.42 × 102 copies of positive control plasmid [Fig. 2A] or as little as 10-2 ng S. japonicum genomic DNA [Fig. 2B] when considering 35 cycles as the cut-off detection limit. No fluorescence signal was obtained when purified DNA from the following was tested: non-infected O. nosophora snails, non-infected mouse faeces, O. viverrini, Centrocestus spp, H. taichui, F. gigantica, E. malayanum, P. heterotremus, Haplorchoides spp, Stellantchasmus spp, animal schistosomes, S. mekongi and S. mekongi-infected mouse faeces, human leukocytes, negative human faeces and human faeces infected with S. stercoralis, Taenia spp, Echinostoma and intestinal lecithodendriid flukes With regards to the limit of detection, as little as a single egg could be detected in 100 mg non-infected mouse faeces based on the 192-bp band in the ethidium bromide-stained gel [Fig. 3A, Lane 2]. DNA samples from each aliquot of one, five and 10 non-infected O. nosophora snail tissues artificially inoculated with one S. japonicum cercaria were amplified [Fig. 3B, Lanes 7-9]. Real-time FRET PCR for the detection of S. japonicum in infected O. nosophora snails and mice - A total of 30 S. japonicum-infected, 30 non-infected O. nosophora snails, 10 S. japonicum-infected and 10 non-infected mouse faecal samples were analysed separately. The melting curve analyses are shown in Fig. 4. When using S. japonicum-specific primers and probes, the mean ± standard deviation (SD), range and the median of the Tm values of the S. japonicum-infected O. nosophora snails were 68.98 ± 0.05, 68.93-69.06 and 68.99, respectively, and those of the S. japonicum-infected mouse faecal samples were 68.64 ± 0.29, 68.12-69.00 and 68.68, respectively. A total of 30 S. japonicum-infected O. nosophora snails (Cn range = 9.61-15.07; mean ± SD = 10.97 ± 2.33; median = 9.84) and 10 S. japonicum-infected mouse faecal samples (Cn range = 22.04-30.03; mean ± SD = 25.41 ± 2.5; median = 25.78) were positive by real-time FRET PCR with melting curve analysis, whereas all of the specific control DNAs were negative. However, no significant correlation between the Cn and the S. japonicum eggs intensity in the faecal samples were reported (p > 0.050) (data not shown). The positive and negative rates were 100%. To ensure this procedure can detect S. japonicum DNA from different strains, DNA extracted from S. japonicum Philippine strain adults and from O. quadrasi snails experimentally infected with the S. japonicum Philippine strain were also examined by this method. All samples were positive and the nucleotide sequences of the amplified products were identical to that of the S. japonicum Japanese Yamanashi strain (data not shown). The validity of the real-time FRET PCR method used in this study was verified by the presence of a prominent 192-bp product, which was amplified from the DNAs of the S. japonicum-infected O. nosophora snails, S. japonicum-infected mouse faeces, S. japonicum cercariae, S. japonicum genomic DNA and the positive control plasmid [Fig. 5, Lanes 1-6, P]. However, non-specific bands were also amplified using genomic DNA from some control materials. Although the genomic DNAs from non-infected mouse faeces [Fig. 5, Lane 8], Centrocestus spp [Fig. 5, Lane 10], H. taichui [Fig. 5, Lane 11], F. gigantica [Fig. 5, Lane 12], Stellantchasmus spp [Fig. 5, Lane 16], animal schistosomes [Fig. 5, Lane 17], intestinal lecithodendriid flukes [Fig. 5, Lane 20], S. mekongi [Fig. 5, Lane 24] and S. mekongi-infected mouse faeces [Fig. 5, Lane 25] demonstrated various amplified bands, no specific fluorescence signal was detected by the melting curve analysis. DISCUSSION The strategies of transmission control and the interruption of schistosomiasis japonicum require sensitive and specific tests, particularly in areas where the level of endemicity decreases (Lier et al. 2009). Real-time PCR has increasingly superseded c-PCR due to its greatly improved molecular detection efficacy. Not only is this method accurate, rapid and can measure the specific DNA quantity in samples (Lyon & Wittwer 2009), it also discriminates the species or strains of several pathogenic agents by melting curve analysis. Furthermore, this method offers a high throughput and is performed in a closed system, which eliminates the risk of potential cross-over contamination because it does not require agarose gel electrophoresis to visualise the amplicons. The real-time FRET PCR-based method has been successfully used for the detection of several parasites, including Wuchereria bancrofti (Lulitanond et al. 2004), Brugia malayi (Thanchomnang et al. 2008, 2010a), Dirofilaria immitis (Thanchomnang et al. 2010b), Toxoplasma gondii (Brenier-Pinchart et al. 2007), Plasmodium falciparum (Ojurongbe et al. 2007) and O. viverrini (Intapan et al. 2008a, b, 2009b). In this report, we demonstrate that real-time FRET PCR combined with melting curve analysis using two individually labelled hybridisation probes can be used for the sensitive and specific detection of S. japonicum infection in snails and faecal samples. This procedure could detect as little as one cercaria implanted in a pool of 10 non-infected O. nosophora snails and a single egg inoculated in 100 mg of non-infected mouse faeces. The results were positive in all S. japonicum-infected snails and faecal samples from infected mice equalling a 100% positive rate. Regarding the analytical specificity of the procedure, no fluorescence was observed despite the amplification of non-specific bands when DNA from some flukes belonging to a different species than S. japonicum was tested, indicating a 100% negative rate. Thus, our real-time FRET PCR could be useful for the differentiation of S. japonicum cercariae from those of other flukes, such as S. mekongi, O. viverrini, Centrocestus spp, H. taichui, F. gigantica, E. malayanum, P. heterotremus, Haplorchoides spp, Stellantchasmus spp and animal schistosomes. This method could also distinguish S. japonicum eggs in mouse faecal samples from DNA extracted from human leukocytes, negative human faeces and human faeces infected with S. stercoralis, Taenia spp, intestinal lecithodendriid flukes and Echinostoma. The real-time FRET PCR protocol provides an alternative to the classic or more modern molecular or serological methods for the detection of S. japonicum in snails, the first intermediate host and in faecal samples. Because the probes and assay system can be used to detect at least two strains of S. japonicum (Japanese and Philippine strains), this method can provide standardised information about S. japonicum epidemiology. The entire protocol (after the extraction of specimen DNA) can be completed within 1 h and thus is high throughput. A large number of samples can be processed simultaneously, and only small sample volumes are required. This method eliminates the need for laborious, time-consuming microscopic examination by experienced personnel. Moreover, it is independent of the subjective bias that is sometimes present in microscopic examinations and avoids confusion with other miscellaneous parasite eggs. Nevertheless, no significant correlation between Cn and the S. japonicum egg amount in the faecal samples was observed. This may be due to miracidium DNA degradation in S. japonicum eggs in the faecal samples. Further studies are needed to improve the quantitative efficacy of this method. In conclusion, a specific, sensitive and fast real-time FRET PCR for the detection of S. japonicum in snail intermediate hosts and in infected mouse faecal samples is reported here. This test is suitable for not only epidemiological studies and eradication programs for intermediate hosts, but also for the possible diagnosis of human infection in Asian populations. ACKNOWLEDGEMENTS To Dr Yukifumi Nawa, for comments and suggestions, and to Dr Mark Roselieb, for his assistance in preparing the paper. REFERENCES
Received 15 March
2011 Copyright 2011 - Memorias do Instituto Oswaldo Cruz The following images related to this document are available:Photo images[oc11170f3.jpg] [oc11170f4.jpg] [oc11170f2.jpg] [oc11170f1.jpg] [oc11170f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}