 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz, Vol. 106, No. 7, November 2011 Hybridism between Biomphalaria cousini and Biomphalaria amazonica and its susceptibility to Schistosoma mansoni Tatiana Maria TeodoroI*; Liana Konovaloff Jannotti-PassosII; Omar dos Santos CarvalhoI; Mario J GrijalvaIII, IV; Esteban Guilhermo BaúsIV; Roberta Lima CaldeiraI ILaboratório
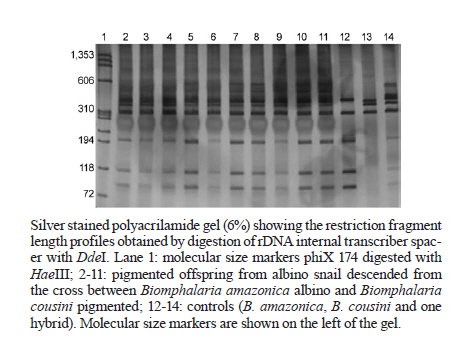
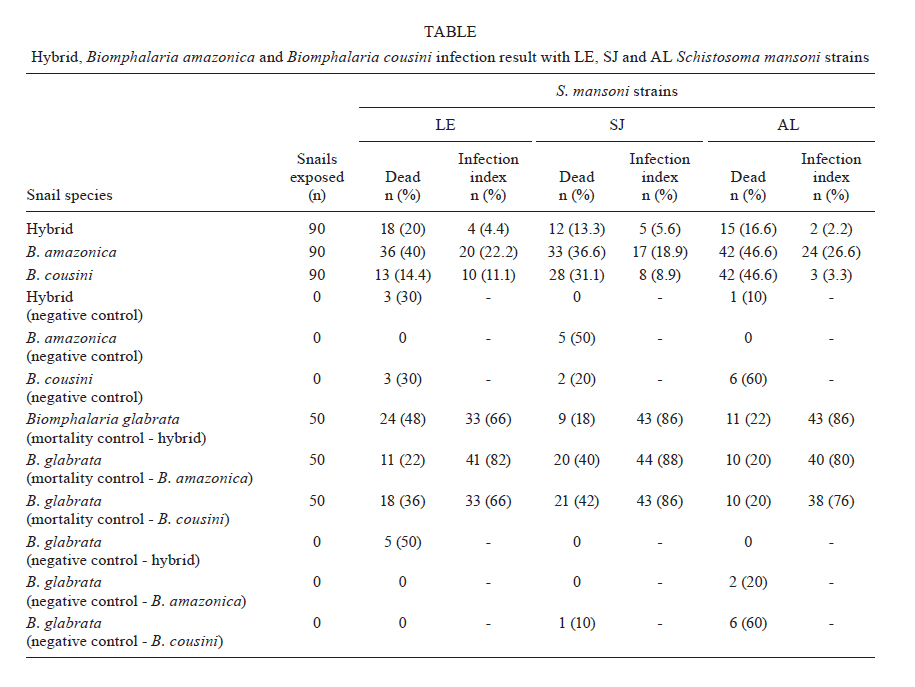
de Helmintologia e Malacologia Médica Code Number: oc11173 ABSTRACT Molecular techniques can aid in the classification of Biomphalaria species because morphological differentiation between these species is difficult. Previous studies using phylogeny, morphological and molecular taxonomy showed that some populations studied were Biomphalaria cousini instead of Biomphalaria amazonica. Three different molecular profiles were observed that enabled the separation of B. amazonica from B. cousini. The third profile showed an association between the two and suggested the possibility of hybrids between them. Therefore, the aim of this work was to investigate the hybridism between B. cousini and B. amazonica and to verify if the hybrids are susceptible to Schistosoma mansoni. Crosses using the albinism factor as a genetic marker were performed, with pigmented B. cousini and albino B. amazonica snails identified by polymerase chain reaction-restriction fragment length polymorphism. This procedure was conducted using B. cousini and B. amazonica of the type locality accordingly to Paraense, 1966. In addition, susceptibility studies were performed using snails obtained from the crosses (hybrids) and three S. mansoni strains (LE, SJ, AL). The crosses between B. amazonica and B. cousini confirmed the occurrence of hybrids. Moreover, hybrids can be considered potential hosts of S. mansoni because they are susceptible to LE, SJ and AL strains (4.4%, 5.6% and 2.2%, respectively). These results indicate that there is a risk of introducing schistosomiasis mansoni into new areas. Key words: Biomphalaria cousini - Biomphalaria amazonica - hybrids - susceptibility - Schistosoma mansoni - Brazil Geographical distribution of the trematode Schistosoma mansoni Sambon, 1907, the etiological agent of intestinal schistosomiasis, is directly associated with the presence of its intermediate hosts, hermaphroditic freshwater snails of the genus Biomphalaria (Gastropoda: Planorbidae). However, not all Biomphalaria species are susceptible to S. mansoni. There are 37 species and one sub-species of molluscs of the genus Biomphalaria, of which nine are susceptible to S. mansoni (Biomphalaria glabrata, Biomphalaria pfeifferi, Biomphalaria alexandrina, Biomphalaria sudanica, Biomphalaria straminea, Biomphalaria tenagophila, Biomphalaria choanomphala, Biomphalaria camerunensis, Biomphalaria prona), nine are potential intermediate hosts (Biomphalaria amazonica, Biomphalaria peregrina, Biomphalaria amazonica aff. straminea, Biomphalaria havanensis, Biomphalaria helophila, Biomphalaria stanley, Biomphalaria sericea, Biomphalaria smithi, Biomphalaria cousini), nine are refractory (Biomphalaria obstructa, Biomphalaria intermedia, Biomphalaria occidentalis, Biomphalaria oligoza, Biomphalaria amazonica tenagophila guaibensis, Biomphalaria kuhniana, Biomphalaria schrammi, Biomphalaria orbignyi, Biomphalaria trigyra) and there is no information as to the status of susceptibility to S. mansoni of the other species (Carvalho et al. 2008, Teodoro et al. 2010). Although morphological studies of Biomphalaria for species identification have been carried out, differentiation between some species may be complicated. The identification of these molluscs is based on anatomical and morphological characteristics and relies on descriptions by Paraense (1966, 1975, 1981, 1984, 1988). Currently, the most important tool for specific identification of Biomphalaria snails is the researcher's capability to dissect and characterise snails based on their morphological differences. The difficulties involved in specific identification based on morphological characters have motivated the use of molecular techniques (Vidigal et al. 2000). Therefore, the correct identification of these snails is of great importance because it enables the detection of species in areas where schistosomiasis transmission occurs, as well as in areas without the disease but that might become schistosomiasis foci owing to the presence of natural or experimentally susceptible species. Studies using Biomphalaria snails found three variant molecular profiles for B. amazonica (Vidigal et al. 2000). Later, studies using phylogeny, morphological and molecular taxonomy of Brazilian, Bolivian and Colombian snails previously identified as B. amazonica showed that some populations studied were B. cousini and established the occurrence of B. cousini in Brazil. Three different molecular profiles were observed, enabling the differentiation of B. amazonica from B. cousini. The third profile showed associations with the two B. cousini and B. amazonica profiles, suggesting the possibility of hybrids between these species (Caldeira et al. 2010, Teodoro et al. 2010). Experiments on susceptibility have shown that B. amazonica and B. cousini are potential hosts of S. mansoni (Corrêa & Paraense 1971, Paraense & Corrêa 1985, Fernandez & Thiengo 2006, Teodoro et al. 2010). Experiments based on crosses of molluscs of the genus Biomphalaria have been performed since the 1950s, using the albinism factor as a genetic marker, which is determined by a pair of recessive alleles (Paraense 1955, 1956, Paraense & Deslandes 1955). Crosses between B. glabrata and B. tenagophila were not feasible reporting the reproductive isolation of these species (Paraense & Deslandes 1955). Nevertheless, crosses between Biomphalaria species that produce fertile hybrids have been reported (Barbosa et al. 1956, 1958, Mello-Silva et al. 1998). In this context, the aim of this work was to investigate the presence of hybrids between B. cousini from the type locality Santo Domingo de Los Colorados, Ecuador and B. amazonica from the type locality Igarape da Cachoeirinha, Manaus, state of Amazonas (AM), Brazil and to verify if these species are susceptible to S. mansoni. MATERIALS AND METHODS Snail populations - The molluscs used for crosses experiments were from type localities defined by Paraense (1966). B. cousini snails were collected from Santo Domingo de Los Colorados (0024303S 07913934W). B. amazonica snails were collected from Igarape da Cachoeirinha (3073274S 60003261W). The molluscs used for susceptibility experiments were from Benjamin Constant, AM (B. cousini and B. amazonica), which exhibited the same profile as the snails from the type locality Belo Horizonte, Minas Gerais (MG), Brazil (B. glabrata) and descended from the cross between B. amazonica and B. cousini from Benjamin Constant (hybrids). The specimens were identified by polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) using the internal transcribed spacer (ITS) region of the rDNA with the primers ETTS2 (5'TAACAAGGTTTCCGTAGGTGAA3') and ETTS1 (5'TGCTTAAGTTCAGCGGGT3'), which were anchored in the conserved extremities of the 18S and 28S ribosomal genes, respectively (Kane & Rollinson 1994). DdeI enzyme was used for each digestion reaction. The snails were maintained in the Lobato Paraense Mollusc Rearing of René Rachou Research Institute/Oswaldo Cruz Foundation under appropriate environmental conditions and were fed lettuce leaves and calcium carbonate. Cross trials - Populations of B. cousini and B. amazonica that were kept in the Lobato Paraense Mollusc Rearing and had been previously identified by PCR-RFLP were used. Using albinism as the genetic marker, five crosses were performed using albino B. amazonica and pigmented B. cousini snails as described below. One adult albino B. amazonica and one pigmented B. cousini were kept separate until they had laid eggs. After hatching, 10 offspring from each species were identified by PCR-RFLP and were kept isolated until they had laid eggs for the first time. Five albino B. amazonica and five pigmented B. cousini snails were selected for the crosses. Each couple was kept in the same container for one week and then was separated. Ten pigmented offspring obtained from each albino snail were individually identified by PCR-RFLP to verify the profile of these descendents. Susceptibility experiments - A total of 90 hybrid specimens (3-5 mm in diameter) were exposed to S. mansoni miracidia of the strain LE (isolated from a patient from Belo Horizonte and kept in the laboratory since 1968). The snails were individually exposed to eight miracidia. Based on previous studies on susceptibility performed with B. amazonica (Corrêa & Paraense 1971, Paraense & Corrêa 1985, Fernandez & Thiengo 2006), a pilot project was conducted using the average number of miracidia used in these papers and the best result was found using eight miracidia, with more infected snails and less mortality. Fifty specimens of B. glabrata were exposed to eight miracidia as an infection control. Ten specimens of B. glabrata and 10 hybrid specimens were used as non-infected controls. Susceptibility studies were performed as described by Pellegrino and Katz (1968) and modified as in Jannotti-Passos et al. (2008). On the 30th day after exposure and subsequently every week, the snails were placed singly in vials with water and exposed to artificial light (28-30ºC) to induce shedding of the cercariae. If any specimen died, it was examined by low stringency-PCR to detect the presence of S. mansoni (Jannotti-Passos et al. 1997). The specimens that survived for 80 days after exposure without shedding cercariae were placed between glass slides and then examined for schistosome development stages. The specimens that shed cercariae were exposed to light for 2 h on alternative days until they died and Swiss albino mice were inoculated with the cercariae to verify their viability. This procedure was repeated with strains SJ (isolated from B. tenagophila in São José dos Campos, state of São Paulo, Brazil and kept in the laboratory since 1975) and AL (isolated from B. glabrata in the state of Alagoas, Brazil and kept in the laboratory since 1980). All the procedures described above were performed using B. amazonica and B. cousini to allow for comparisons between them. RESULTS Crossing trials - The PCR amplification of the Biomphalaria ITS region with the primers ETTS2 and ETTS1 resulted in a product of approximately 1.3 kb (data not shown). The RFLP profiles obtained by digesting rDNA ITS with DdeI are represented in Figure, with the profile of the albino snail corresponding to the profile of B. amazonica (Lane 12) and the pigmented snail corresponding to B. cousini (Lane 13). The profiles of the 10 pigmented offspring obtained from each albino snail correspond to the profile of the hybrid control (Lane 14). The five crosses generated the same result and only one gel is represented. Susceptibility experiments - The hybrids were susceptible to the LE, SJ and AL strains of S. mansoni, (4.4%, 5.6% and 2.2%, respectively). All infected snails shed cercariae, which were determined to be viable by mouse inoculation. B. amazonica and B. cousini were also susceptible to the LE, SJ and AL strains of S. mansoni (22.2%, 18.9% and 26.6% to B. amazonica and 11.1%, 8.9% and 3.3% to B. cousini, respectively) [Table]. DISCUSSION Previous studies have suggested the possibility of natural hybrids between B. amazonica and B. cousini (Teodoro et al. 2010). Our results leave no doubt about the possibility of a hybrid between B. amazonica and B. cousini, which we confirmed through a cross between these species using the albinism factor as a genetic marker (determined by a pair of recessive alleles), as standardised to Biomphalaria by Paraense (1955). Experiments based on crosses of molluscs of the genus Biomphalaria have been performed since the 1950s (Paraense 1955, 1956, Paraense & Deslandes 1955). Crosses between Biomphalaria species that produced fertile hybrids have been reported. Examples include crosses between B. alexandrina and B. glabrata (Barbosa et al. 1956): B. straminea and B. peregrine, B. tenagophila and B. peregrina (Barbosa et al. 1958) and B. glabrata and B. tenagophila (Mello-Silva et al. 1998). Barbosa (1964) commented on the natural hybridisation between B. glabrata and B. tenagophila, which generates hybrids with intermediate forms. Subsequently, Barbosa (1973) described observations made over three years on the replacement of B. glabrata by B. straminea in northeastern Brazil, where four specimens that he called interspecific hybrids were found. Only during this process of competitive exclusion were mixed forms found, possibly as a result of crosses between these species. Later, new field studies were conducted and these intermediate forms were not found, revealing their inability to persist in nature. Confirming our speculations, the pigmented descendents obtained from each albino snail had the hybrid profile [Figure]. Thus, this study confirms that these two species produce hybrids. Mello-Silva et al. (1998) commented that the results of fertility and fecundity tests of fertile hybrids revealed that the F2 generation was not as efficient as the F1, involving the formation of normal, vigorous and fertile F1 generation hybrids, but a weak F2 generation with the presence of some sterile individuals. However, further studies are necessary to better understand the biology of these hybrids. Once the population of hybrids was defined, we examined their susceptibility to S. mansoni. Some species of Biomphalaria have shown different susceptibilities to infection by S. mansoni (Paraense & Corrêa 1963, 1973, 1978, 1985). Experiments on susceptibility have shown that B. amazonica and B. cousini are potential hosts of S. mansoni (Corrêa & Paraense 1971, Paraense & Corrêa 1985, Fernandez & Thiengo 2006, Teodoro et al. 2010). Specimens of B. amazonica from Careiro da Várzea (AM) exhibited a higher infection index to two strains of S. mansoni (48% and 73%) (Corrêa & Paraense 1971). A study using B. amazonica from Porto Velho (state of Rondônia) showed a lesser degree of compatibility between the strain and the snail used (3.5%) (Paraense & Corrêa 1985). Fernandez and Thiengo (2006) confirmed this result using B. amazonica from the state of Mato Grosso, with a low index of infection (3.79%, 13.63% and 9.09%). Teodoro et al. (2010) showed a higher infection index of B. cousini to the LE strain of S. mansoni (31.6%). Our results showed that hybrids can be considered a potential host of the trematode because they proved to be susceptible to the three strains of S. mansoni used, LE, SJ and AL (4.4%, 5.6% and 2.2%, respectively). Confirming the previous results obtained with B. amazonica and B. cousini, these species were shown to be susceptible to the LE, SJ and AL strains of S. mansoni (22.2%, 18.9% and 26.6%, respectively, to B. amazonica, and 11.1%, 8.9% and 3.3%, respectively, to B. cousini). Nevertheless, high mortality was observed for B. amazonica when compared to B. cousini and hybrid snails. However, a higher index infection was observed for B. amazonica when compared to hybrids and B. cousini. This result can be related to the poor adjustment between B. cousini and hybrids snails to the strains used being B. amazonica more compatible with the strains used. Additionally, the susceptibility of Biomphalaria snails to infection by S. mansoni is a genetically controlled and heritable trait over generations (Newton 1953). According to Lewis et al. (1993), the host/parasite interactions are influenced by mollusc genes that control susceptibility and parasite genes that determine infectivity. This condition establishes different levels of host/parasite interactions. Mortality was observed in the snails used as the negative control. This relatively short survival can be attributed to the long period of experimentation and to adjustments to the artificial environment. Thus, the identification of Biomphalaria snails is of great importance because the presence of the species (B. amazonica, B. cousini and hybrids between than) may result in schistosomiasis foci because these species are experimentally susceptible to S. mansoni. ACKNOWLEDGEMENTS To the Lobato Paraense Mollusc Rearing of CPqRR/Fiocruz, to the support for this research, and to José Geraldo Amorim da Silva and Jairo, CPqRR/Fiocruz, for technical support. REFERENCES
Received 20 April
2011 Copyright 2011 - Memorias do Instituto Oswaldo Cruz The following images related to this document are available:Photo images[oc11173f1.jpg] [oc11173t1.jpg] |
| |||||||||

{kind=link}
{kind=link}