 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
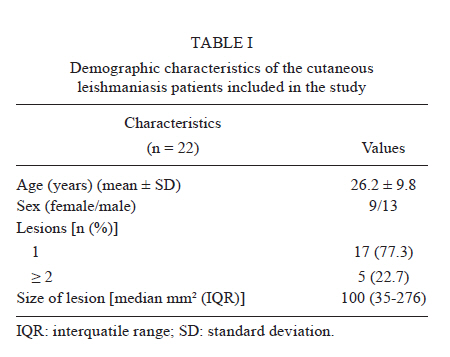
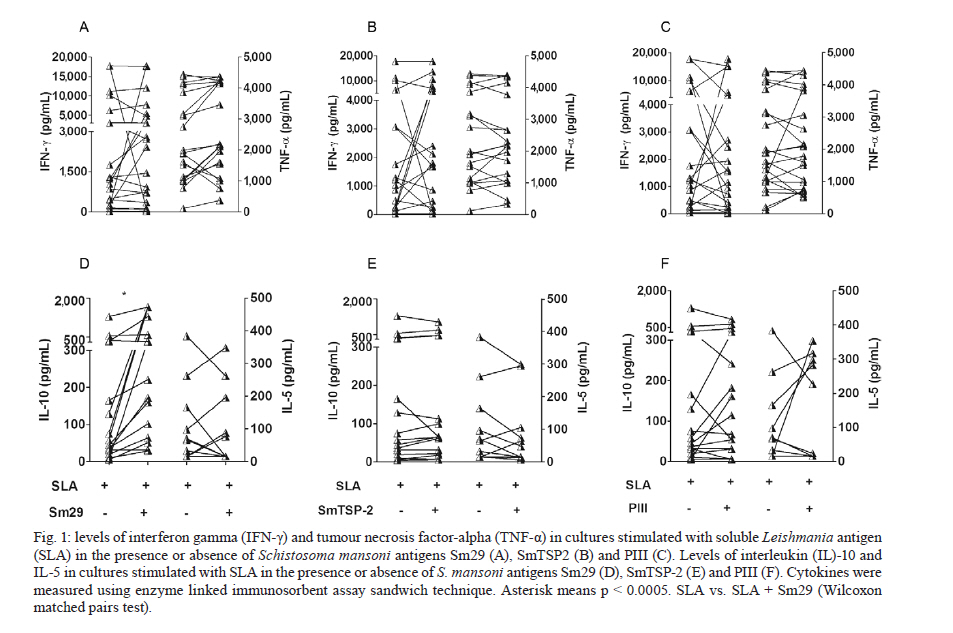
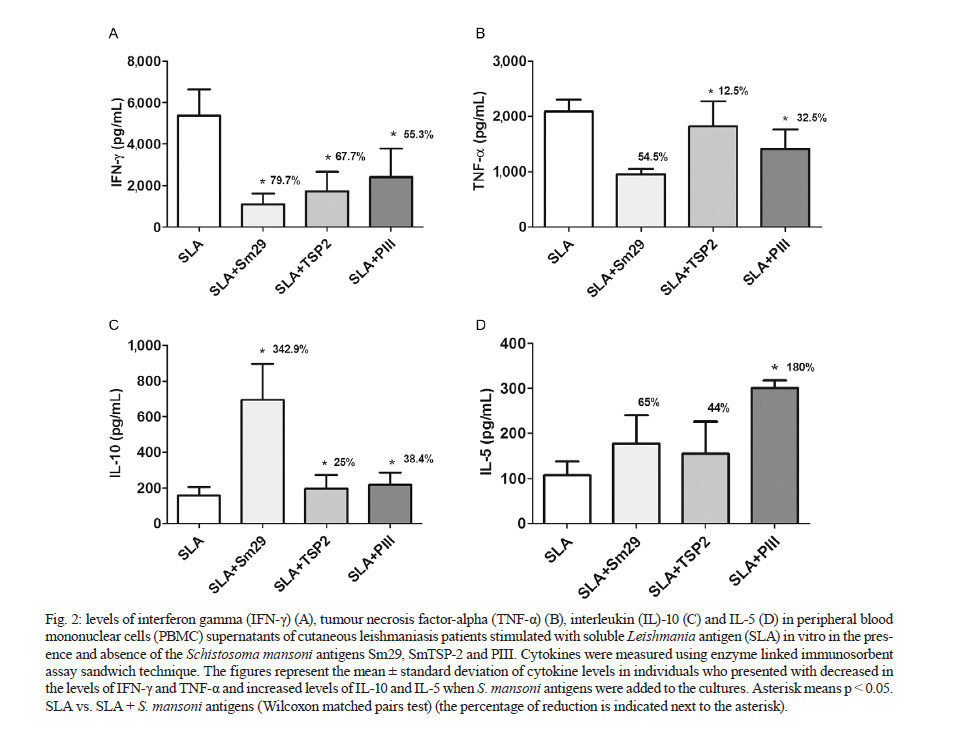
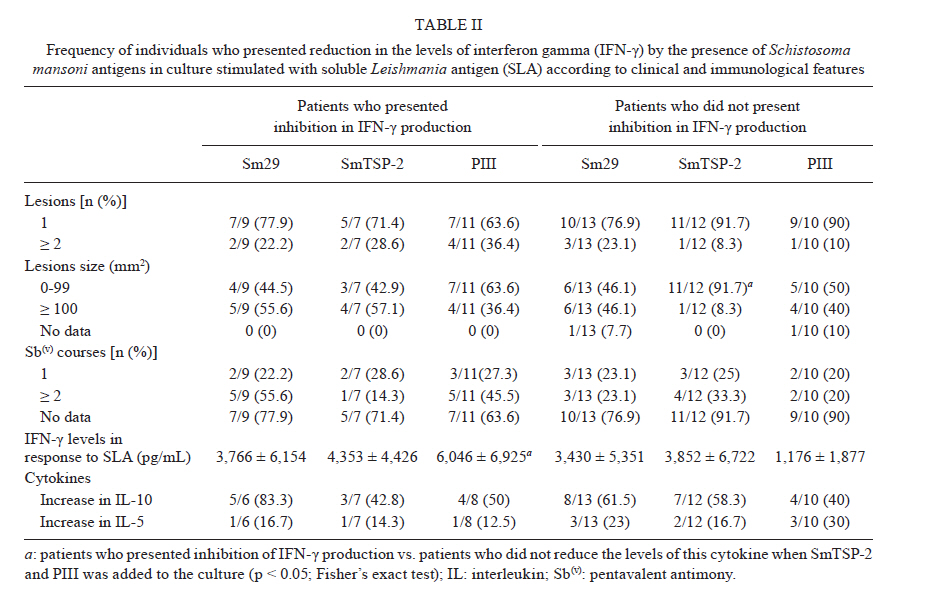
Memorias do Instituto Oswaldo Cruz, Vol. 106, No. 7, November 2011 Schistosoma mansoni antigens alter the cytokine response in vitro during cutaneous leishmaniasis Aline Michelle Barbosa BaficaI*; Luciana Santos CardosoI, II; Sérgio Costa OliveiraIII; Alex LoukasIV; Giuseppe Tittoni VarelaI; Ricardo Riccio OliveiraI; Olívia BacellarI, V; Edgar Marcelino CarvalhoI, V, VI; Maria Ilma AraújoI, V, VI IServiço
de Imunologia, Hospital Universitário Professor Edgard Santos, Universidade
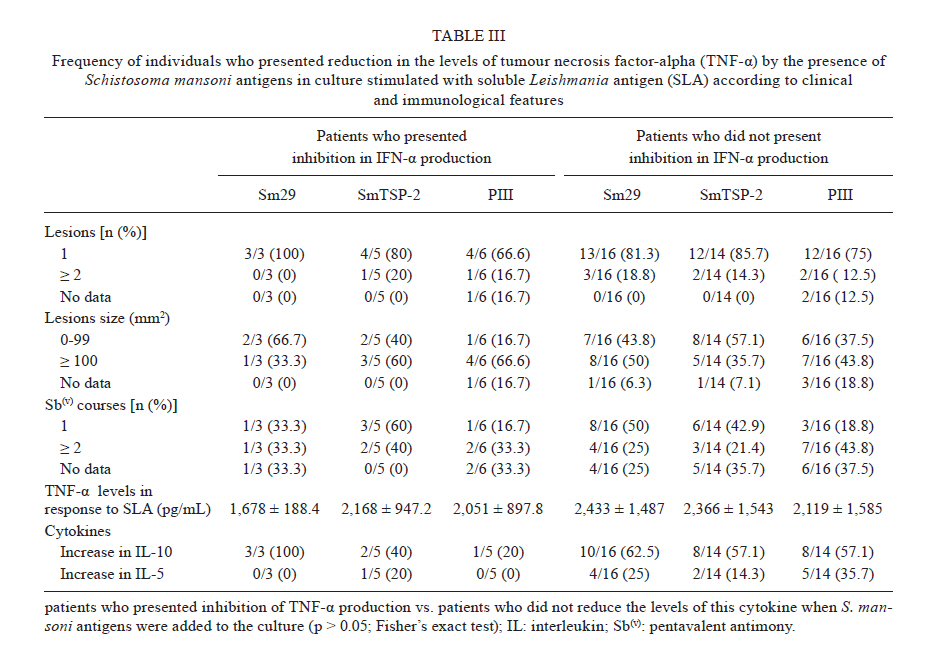
Federal da Bahia, Rua João das Botas s/n, 40110-160 Salvador, BA, Brasil Code Number: oc11174 ABSTRACT Schistosoma mansoni infection or associated products are able to down-modulate the type 1 CD4+ T cell inflammatory response characteristic of autoimmune diseases. In this study, we evaluated how S. mansoni antigens altered the immune response that was induced by the soluble Leishmania antigen (SLA) from cutaneous leishmaniasis (CL) patients. Cytokines were measured from the supernatants of peripheral blood mononuclear cell cultures stimulated with SLA. This was performed using the sandwich enzyme linked immunosorbent assay technique in the presence or absence of S. mansoni recombinant antigens Sm29, SmTSP-2 and PIII. The addition of S. mansoni antigens to the cultures resulted in the reduction of interferon gamma (IFN-γ) levels in 37-50% of patients. Although to a lesser extent, the antigens were also able to decrease the production of tumour necrosis factor-alpha (TNF-α). We compared patients that either had or did not have reduction in IFN-γ and TNF-α production in cultures stimulated with SLA in the presence of S. mansoni antigens. We found that there was no significant difference in the levels of interleukin (IL)-10 and IL-5 in response to S. mansoni antigens between the groups. The antigens used in this study down-modulated the in vitro proinflammatory response induced by SLA in a group of CL patients through a currently undefined mechanism. Key words: cutaneous leishmaniasis - S. mansoni antigens - Sm29 - SmTSP-2 - PIII Leishmaniasis is an infectious disease caused by a protozoan from the genus Leishmania spp. It is one of the most common infectious diseases, affecting 12 million people worldwide with an incidence of 1.5 million cases per year (Desjeux 2001). Four different forms of tegumentary leishmaniasis are described: cutaneous, mucosal, diffuse and disseminated leishmaniasis. Cutaneous leishmaniasis (CL) is the most common clinical manifestation, characterised by one to several skin lesions in exposed areas, with an absence or small number of parasites (Jones et al. 1987, Bogdan et al. 1996). CL is endemic in many regions of Central and South America (Jones et al. 1987). In the state of Bahia (BA), in Brazil, the number of CL cases reaches 23.4 per 100,000 inhabitants (SVS/MS 2010). Some evidence has suggested that the immune response contributes to the tissue injury characteristic of CL. Leishmania donovani infection is controlled by type 1 CD4+ T cell (Th1) immune response. The Th1 response is important to produce interferon-gamma (IFN-γ) and activate tissue macrophages (Murray & Cartelli 1983, Murray et al. 1983). However in the presence of a strong of a Th1 response, there is an increased production of IFN-γ and tumour necrosis factor-alpha (TNF-α). This increased production of inflammatory cytokines leads to intense tissue damage resulting in the development of CL and mucosal leishmaniasis (Ribeiro-de-Jesus et al. 1998). Bacellar et al. (2002) showed that mononuclear cells from mucosal leishmaniasis patients have a decreased ability to produce and to respond to interleukin (IL)-10 after restimulation with Leishmania braziliensis antigen in vitro. Moreover, neutralisation of IL-10 enhanced IFN-γ production by peripheral blood mononuclear cells (PBMC) and also increased lymphocyte proliferation in patients with CL (Rocha et al. 1999). The regulatory cytokine IL-10 seems to play a major role in modulating the inflammatory responses associated with CL development. However, this cytokine maintains latency in murine models of Leishmania major infection (Belkaid et al. 2001). In recent years, studies have demonstrated that helminth infections or products from their infections have the potential to modulate Th2-immune responses that result in the pathology of allergic diseases (Araujo et al. 2000, 2004, Cooper et al. 2003). Our research group has demonstrated that asthmatic patients who are infected with Schistosoma mansoni have a less severe course of asthma and an inhibition of the Th2 inflammatory response that seems to be mediated by IL-10 (Medeiros et al. 2004, Cardoso et al. 2006). In a murine model of ovalbumin (OVA)-induced airway inflammation, S. mansoni antigens reduced the allergic Th2 response (Cardoso et al. 2010). Other recent experimental studies have shown that S. mansoni infection or parasite products induce regulatory cells and cytokines that are able to prevent an autoimmune Th1 inflammatory response seen in diseases such as type-I diabetes, encephalomyelitis and psoriasis (Cooke et al. 1999, Sewell et al. 2003, Atochina & Harn 2006). Considering the demonstrated ability of S. mansoni antigens to prevent some Th1-mediated diseases, this study aimed to evaluate the effects of S. mansoni antigens on the immune response induced by the soluble Leishmania antigen (SLA) in CL patients. We examined IFN-γ, TNF-α, IL-10 and IL-5 production by PBMC stimulated in vitro with SLA in the presence or absence of the S. mansoni antigens Sm29, tetraspanin 2 (SmTSP-2) and PIII, a fraction of S. mansoni soluble adult worm antigen (SWAP). PATIENTS, MATERIALS AND METHODS This study evaluated the ability of S. mansoni antigens to alter cytokine production by PBMC from patients with CL, which was assessed based on the PBMC cytokine response to SLA in vitro. The study included 22 individuals living in the endemic area of Corte de Pedra, BA. They attended the local healthcare facility from September 2009-October 2010 and agreed to participate in this study. All patients included in the study donated blood for PBMC cultures, which were stimulated with SLA in the presence or absence of S. mansoni antigens. S. mansoni antigens - The antigens used in this study included the recombinant proteins Sm29 and SmTSP-2 and a fraction of S. mansoni SWAP obtained by anionic chromatography referred to in this study as PIII. The recombinant proteins were cloned in Escherichia coli. They were tested for lipopolysaccharide (LPS) contamination using a commercially available Chromogenic LAL Kit (Cambrex). The levels of LPS in Sm29 and SmTSP-2 were below 0.25 ng/mL. In order to neutralize the potential effects of LPS, polymyxin B was added to the cell cultures every 12 h as previously performed (Cardoso et al. 2007). PBMC cultures and cytokine measurements - PBMCs were obtained using a Ficoll-Hypaque gradient. PBMCs were cultured at a concentration of 3 x 106 cells/mL in Roswell Park Memorial Institute 1640 containing 10% normal human serum (AB+, heat inactivated), 100 U/mL penicillin, 100 mg/mL streptomycin, 2mM L-glutamine and 30 mM HEPES (Life Technologies Gibco-BRL, Gaithersburg, MD). Cells were cultured with the antigens Sm29, SmTSP-2 and PIII at a concentration of 5 µg/mL in the presence or absence of SLA (5 µg/mL). The levels of IFN-γ, TNF-α, IL-10 and IL-5 were measured in the supernatants of PBMC cultures as determined by sandwich enzyme linked immunosorbent assay. The results are expressed as picograms per millilitre (pg/mL) based on a standard curve. Statistical analyses - Statistics were analysed utilising the software Statistical Package for Social Science version 9.0 for Windows. Statistical differences between the means of cytokine levels were analysed using Wilcoxon matched pairs test or Mann-Whitney test, as indicated. Fisher's exact test was used to compare proportions. Statistical significance was established at the 95% confidence interval. The Ethical Committee of Climério de Oliveira Maternity, Federal University of Bahia, approved the present study. Informed consent was obtained from all study participants or their legal guardians. RESULTS The demographic information for CL patients included in the study is shown in Table I. A total of 22 patients with CL were enrolled in this study: 13 were male and nine were female, with a mean age of 26.2 ± 9.8 years (range 6-48 years) [Table I]. The majority of patients presented with a single lesion (77.3%) and the median lesions size was 100 mm² (interquartile range, 35.0-276.0). To test the ability of S. mansoni antigens, Sm29, SmTSP-2 and PIII, to down-modulate IFN-γ and TNF-α production, these antigens were added into SLA-stimulated PBMC cultures from CL patients. There were no significant differences between the mean levels of IFN-γ detected in supernatants of PBMC cultures stimulated with SLA alone [mean ± standard error (SEM) = 3,568 ± 1,184 pg/mL] or SLA plus S. mansoni antigens rSm29, SmTSP-2 and PIII (3,819 ± 1,144 pg/mL, 4,822 ± 1,373 pg/mL and 3,466 ± 308.5 pg/mL, respectively) [Fig. 1A-C]. Likewise, no difference was observed in levels of TNF-α in the supernatants of PBMC cultures stimulated with SLA alone (2,314 ± 318.5 pg/mL) and SLA plus S. mansoni antigens Sm29, SmTSP-2 and PIII (2,638 ± 325.5 pg/mL, 2,314 ± 381.5 pg/mL and 2,167 ± 304.4 pg/mL, respectively) [Fig. 1A-C]. A significant increase in the concentration of IL-10 was observed when Sm29 was added to the cultures (mean ± SEM = 528.8 ± 151.8 pg/mL) as compared to SLA alone (170.3 ± 70.7 pg/mL, p < 0.0005). The difference in mean levels of IL-10 was not significant for the two other S. mansoni antigens used in this study SmTSP-2 and PIII (175.2 ± 64.6 pg/mL and 172.6 ± 53.4 pg/mL, respectively) [Fig. 1D-F]. Additionally, we assessed IL-5 production and found no significant differences between the mean levels of this cytokine detected in the supernatants of PBMC cultures stimulated with SLA alone (mean ± SEM = 66.4 ± 23 pg/mL) or SLA plus S. mansoni antigens rSm29, SmTSP-2 and PIII (62.5 ± 22.3 pg/mL, 56.7 ± 20.3 pg/mL and 89.7 ± 29.4 pg/mL, respectively, p > 0.05) [Fig. 1D-F]. There was, however, a group of patients who presented with reduced levels of IFN-γ and TNF-α and increased levels of IL-10 and IL-5 when Sm29, SmTSP-2 and PIII were added to the cultures stimulated with SLA. The frequency of individuals with reduced IFN-γ production as a result of the presence of Sm29, SmTSP-2 and PIII were 40.9%, 36.8% and 50%, respectively. Reduction in TNF-α levels was found in 15.8%, 26.3% and 27.3% of CL patients when Sm29, SmTSP-2 and PIII were added to the PBMC cultures stimulated with SLA. The addition of Sm29, SmTSP-2 and PIII to the cultures resulted in increased levels of IL-10 in 68.4%, 52.6% and 47.4% of patients, respectively. Additionally, levels of IL-5 were increased in 21%, 15.8% and 26.3% of patients when Sm29, SmTSP-2 and PIII were added to the cultures stimulated with SLA. Patients that showed varying levels of cytokine production following the addition of the S. mansoni antigens to the cultures stimulated with SLA were further analysed individually. The levels (mean ± SEM) of IFN-γ, TNF-α, IL-10 and IL-5 in the supernatants of PBMC cultures stimulated with SLA in the presence or absence of Sm29, SmTSP-2 and PIII are shown in Fig. 2. The mean levels of IFN-γ [Fig. 2A] diminished from 5,379 ± 1,253 pg/mL to 1,093 ± 540.3 pg/mL (79.7% reduction) to 1,739 ± 920.7 pg/mL (67.7% reduction) and to 2,409 ± 1,357 pg/mL (55.3% reduction) in the presence of Sm29, SmTSP-2 and PIII, respectively, compared to SLA alone (p < 0.05). The levels of TNF-α [Fig. 2B] dropped from 2,090 ± 210.8 pg/mL in PBMC cultures stimulated with SLA to 950.9 ± 96.1 pg/mL (54.5% reduction), 1,823 ± 450.8 pg/mL (12.7% reduction) and 1,411 ± 356 pg/mL (32.5% reduction) in the presence of Sm29, SmTSP-2 and PIII, respectively. On the other hand, the levels of IL-10 [Fig. 2C] increased from 156.6 ± 46.9-693.6 ± 204 pg/mL (342.9% increase after addition of Sm29), to 195.7 ± 77.9 pg/mL (25% increase in response to SmTSP-2) and to 216.7 ± 77.9 pg/mL (38.4% increase in response to PIII) (p < 0.05). There was also an increase in the levels of IL-5 from 107.4 ± 30-177.3 ± 62.9 pg/mL (65% increase) to 154.7 ± 71.2 pg/mL (44% increase) and to 301 ± 16.7 pg/mL (180% increase) when Sm29, SmTSP-2 and PIII were added to the cultures, respectively (p < 0.05) [Fig. 2D]. Correlations between the levels of the pro-inflammatory cytokines IFN-γ and TNF-α with the anti-inflammatory cytokines IL-10 and IL-5 were analysed. There was a positive correlation between the levels of TNF-α and IL-5 when Sm29 (r = 0.6; p = 0.004) and SmTSP2 (r = 0.5; p = 0.004) were added to the cultures. No significant correlation was observed between levels of IFN-γ and IL-5 or IFN-γ and IL-10. Additionally, there was no correlation between the levels of TNF-α and IL-10 when the S. mansoni antigens were added to the cultures (not shown). In order to evaluate if the severity of leishmaniasis, measured by number and size of lesions, interferes with the response to S. mansoni antigens in vitro, we analysed the frequency of CL patients who presented or did not present reduction in the levels of IFN-γ and TNF-α according to these specified parameters [Tables II, III]. Patients whose levels of IFN-γ were unaffected by the addition of SmTSP-2 to the cultures had smaller lesion size compared to patients who had reduced levels of IFN-γ (p < 0.05) [Table II]. There was no significant difference in other assessed clinical parameters, such as number of lesions and number of pentavalent antimony Sb(v) courses, between these two groups of patients when S. mansoni antigens were added to the PBMC cultures. Additionally, patients who either had reduced levels of IFN-γ and TNF-α or not when S. mansoni antigens were added to the cultures were evaluated regarding the levels of IL-10 and IL-5. Patients with reduced IFN-γ production as a result of the addition of S. mansoni antigens and those patients in whom IFN-γ production was unaffected had similar frequencies of individuals who displayed increased levels of IL-10 and IL-5 in response to S. mansoni antigens (p > 0.05) [Table II]. The frequency of patients who had increased levels of IL-10 and IL-5 when the S. mansoni antigens were added to the cultures did not differ between the group of patients who either had a reduction in TNF-α production or not in the presence of S. mansoni antigens (p > 0.05) [Table III]. Finally, we compared the levels of IFN-γ and TNF-α in response to SLA between the group of patients who had reduced levels of these cytokines by the addition of S. mansoni to the cultures and those who did not have reduced cytokine production. The levels of TNF-α were similar between groups (p > 0.05) [Table III]. However, the levels of IFN-γ were higher in patients who had reduced IFN-γ production in response to PIII (6,046 ± 6,925 pg/mL) as compared to those who did not have reduced production of IFN-γ when PIII was added to the culture (1,176 ± 1,877 pg/mL, p = 0.05). DISCUSSION The present study evaluated the ability of S. mansoni antigens to down-modulate the inflammatory response to SLA in PBMC of CL patients. We showed that the addition of the S. mansoni antigens, Sm29, SmTSP-2 and PIII, to PBMC cultures of patients infected with L. braziliensis stimulated with SLA reduced IFN-γ and TNF-α; conversely, these cells produced increased levels of IL-10 as seen in a considerable number of patients. There is evidence that Schistosoma spp infection or its products protect against the development of Th2-mediated diseases in humans and mice (Araujo et al. 2000, 2004, Medeiros et al. 2003, 2004, Pacifico et al. 2009). Moreover, in experimental studies, it has been shown that S. mansoni infection or its parasite products are able to down-modulate the Th1 inflammatory response that is implicated in several autoimmune diseases, such as type-I diabetes, encephalomyelitis and psoriasis (Cooke et al. 1999, Sewell et al. 2003, Atochina & Harn 2006). Our group has performed studies in an attempt to identify S. mansoni antigens with regulatory properties that may enable them to down-regulate inflammation associated with particular immune-mediated diseases. For instance, in experimental models of OVA-induced asthma, the injection of Sm22.6, PIII or Sm29 antigens resulted in a decrease in Th2-inflammatory mediators involved with disease pathology (Cardoso et al. 2010). Mechanisms underlying this modulation appear to include regulatory pathways induced by S. mansoni or their products. Indeed, it has been shown that during chronic S. mansoni infection, innate immune cells, T cells and T regulatory cells are able to produce IL-10 (Araujo et al. 2004, Hesse et al. 2004, Oliveira et al. 2009). It has been demonstrated that S. mansoni phosphatidilserine (PS) has the ability to stimulate antigen-presenting cells from naÏve individuals to produce IL-10 via Toll-like receptor-2 stimulation which promotes T-regulatory cell maturation (van der Kleij et al. 2002). Moreover, it has been demonstrated that the immune response to the Schistosoma haematobium Toll-like ligand antigen, lyso-PS, results in the production of IL-10 by cells of the innate immune system in non-infected children (van der Kleij et al. 2004). This suggests that there are molecular patterns associated with helminths that are involved in the down-regulation of the immune response. There is evidence that an exacerbated Th1 immune response with high production of IFN-γ and TNF-α, as well a lack of regulatory response, is associated with lesion development in CL and mucosal leishmaniasis, as reviewed by Ribeiro-de-Jesus et al. (2008). On the other hand, these cytokines are fundamental to parasite killing (Liew et al. 1990). IL-10, on the other hand, is able to control the exacerbated Th1 inflammatory response, despite data that show it could be responsible for parasite maintenance (Belkaid et al. 2001). An equilibrate immune response may be the key to a harmless host-parasite relationship. Studies of patients with concurrent infections of L. braziliensis and a helminth infection have shown that these patients tend to present with smaller ulcers than patients without helminth infections. However, the time for the lesion to heal was approximately double in co-infected patients (O'Neal et al. 2007). The authors postulate that these effects are due to a stronger Th2-immune response induced by the helminth infection, which adversely affects the Th1 immune response, which compromises host defence against Leishmania sp. infection. However, another study demonstrated that early introduction of anti-helminthic therapy did not improve clinical outcomes in patients co-infected with helminths and L. braziliensis (Newlove et al. 2011). The antigens used in this study were selected because they are secreted by the membrane and/or tegument of the S. mansoni adult worm. Proteins secreted or localised on the surface of Schistosoma spp, which are in intimate contact with host tissues, can be more effective at triggering immunoregulatory processes (Simpson et al. 1990). Our hypothesis is that these antigens may reduce the Th1 response by a mechanism involving regulatory cells and cytokines, rather than a shift towards a Th2 immune response. Since there is limited production of IL-10 in CL (Bacellar et al. 2002), the use of S. mansoni antigens capable of inducing IL-10 production should be of benefit to these patients. Our findings show that the S. mansoni antigens, SmTSP-2 and PIII, were able to reduce IFN-γ and TNF-α production. However, there was not a large increase in the levels of IL-10. These results suggest that IL-10 production is not the only mechanism involved in down-modulation of the inflammatory response to Leishmania infection. Although cells from patients with CL might produce IFN-γ and TNF-α to delay the infection, the modulation of these cytokines at the site of the lesion appears to be a rational strategy to limit the local exacerbated immune response. In the conventional treatment with Sb(v) there is a prolonged healing time of three-four months. This treatment has serious side effects including pancreatitis, liver enzyme abnormalities and cardiac arrhythmia (Berman 1997). The association of immunomodulators, such as granulocyte/macrophage colony stimulating factor locally applied as adjuvant therapy in low doses, improves the healing of chronic ulcers (Jaschke et al. 1999). Other drugs, such as pentoxifylline, inhibit TNF-α synthesis and thus down-modulates the immune response when associated with antimony therapy. This results in an increased cure rate and a decreased healing time of CL and mucosal leishmaniasis (Lessa et al. 2001, Machado et al. 2007). Therefore, the combination of conventional treatment with an immunomodulatory strategy may be more efficient in preventing or diminishing the tissue damage in CL or mucosal leishmaniasis. In this study, we observed that patients who had reduced production of IFN-γ in the presence of the S. mansoni antigens, Sm29 and PIII, show no differences in clinical parameters, such as number and size of lesion. Additionally, there was no difference in the course number of antimony therapy compared to those patients who did not have reduced levels of IFN-γ. Patients who did not have reduced IFN-γ production in the presence of SmTSP-2 had smaller sized lesions compared to those who had reduced IFN-γ production in the presence of this antigen. Nevertheless, there was no significant difference in the clinical parameters when we compared patients with reduced TNF-α production in the presence of the S. mansoni antigens and those who did not have reduced TNF-α production. Additionally, we compared the frequency of patients who had increased levels of IL-10 and IL-5 when S. mansoni antigens were added to the cultures in the groups of patients who either had reduced production of IFN-γ and TNF-α or production was unaffected when cultured with S. mansoni antigens. We found no significant difference between them. These findings indicate that even though S. mansoni antigens are capable of inducing Th2 and T regulatory cytokine production, other regulatory mechanisms may have a role in the down-modulation of Th1-inflammatory response in CL. Alternatively, IL-10 might be essential but not sufficient to regulate IFN-γ and TNF-α production. This concept has been demonstrated where expression of IL-10 receptor was impaired in mucosal leishmaniasis (Faria et al. 2005). This could explain the results in our study where IL-10 did not down-regulate T cell responses in cells isolated from CL patients. It has been shown that inhibition of IFN-γ production tends to be greater at lower concentrations of this cytokine (Ito et al. 1999). In our study, the base levels of IFN-γ in response to SLA were compared between the group of patients who had reduced IFN-γ production with the addition of S. mansoni antigens to the cultures and those patients whose IFN-γ production was unaffected. No significant differences were found in the mean levels of IFN-γ in response to SLA in patients who had reduced levels of IFN-γ with the addition of Sm29 and SmTSP-2 compared to the group who did not have reduced levels of IFN-γ. Unexpectedly, the base levels of IFN-γ were higher in patients who had reduced levels of IFN-γ with the addition of PIII compared to the group who did not have reduced levels of IFN-γ. The mean levels of TNF-α in response to SLA did not differ between patients based on the levels of expression of TNF-α that followed the addition of S. mansoni antigens. These data lead us to conclude that the base levels of pro-inflammatory cytokines exert little or no influence on the immune response to bystander antigens in CL. Taken together, we have shown that the S. mansoni antigens Sm29, SmTSP-2 and PIII induce IL-10 and IL-5 production in CL. Additionally, these antigens are able to control the in vitro inflammatory response in patients independent of the clinical features of disease, such as number and size of lesions. Ongoing studies are being conducted to identify other regulatory mechanisms that may be involved in the down-modulation of the inflammatory response in CL. ACKNOWLEDGEMENTS To Dr Alfredo Goes, for his support in the development of this work, to Michael Sundberg, for his review of the manuscript, and to Dr Luiz Henrique Guimarães, Dr Paulo Machado and Ednaldo Lago, for their assistance in the endemic area. REFERENCES
Received 27 April
2011 Copyright 2011 - Memorias do Instituto Oswaldo Cruz The following images related to this document are available:Photo images[oc11174f1.jpg] [oc11174t3.jpg] [oc11174t2.jpg] [oc11174f2.jpg] [oc11174t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}