 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 89(1): 21-24,Jan./mar. 1994 Ultrastructural Aspects of Virus Replication in One Fatal Case and Several Ortrud Monika Barth, Luzia Monteiro de Castro Crtes, Elisabeth Lampe,Jos da Costa Farias Filho Departamento de Virologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil This work received financial support from the ministerio da saude, Brazil. Received 25th May 1993, Accepted 30th November 1993
Code Number:oc94004 Sizes of Files:
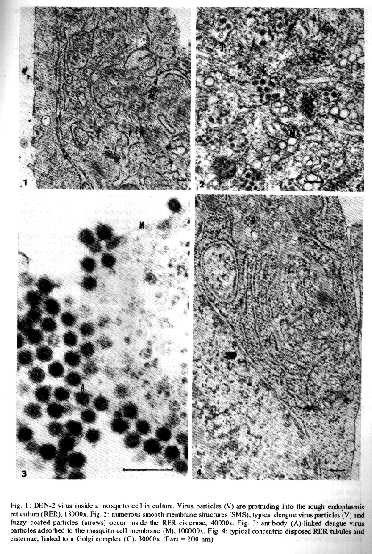
Dengue virus replication in mosquito cell cultures was observed by electron microscopy in one fatal and 40 classical isolates from a dengue type 2 outbreak in Rio de Janeiro and compared with the prototype New Guinea C strain. All the Brazilian isolates presented, beside the classical structured dengue virus particles, fuzzy coated virus-like particles, never observed in the referencial New Guinea C virus strain. More numerous DEN-2 virus particles, fuzzy coated virus-like particles, defective virus particles and smooth membrane structures inside the rough endoplasmic reticulum characterized the unique fatal isolate examined. Key words: Flaviviridae dengue-2 virus replication ultrastructure During the epidemic of dengue serotype 2 (DEN-2) in Rio de Janeiro (Nogueira et al. 1990), following a previous outbreak of dengue serotype 1 (DEN-1) in 1986 (Schatzmayr et al. 1986), some fatal occurrences were noticed. Virus replication was observed by electron microscopy in mosquito cell cultures, inoculated with positive human sera, as well as the occurrence of other structures than DEN-2 virus particles. Virus replication involve the introduction of viral nucleic acids and viral proteins inside the rough endoplasmic reticulum (RER), which further are processed for maturation and cell release (Barth 1992, Barth & Schatzmayr 1992). Only a few virus particles probably reach the infective stage, the majority remaining inside the RER. Portions of the RER develop to smooth membrane vesicles from which the virus particles are never released. In order to get more information about virus replication of different isolates from the DEN-2 epidemic at Rio de Janeiro, light and electron microscope observations were made in comparison to a fatal case, described by Nogueira et al. (1991), from the same occurrence. MATERIALS AND METHODS Viruses The DEN-2 viruses analyzed were obtained from 41 patients during the 1990 dengue epidemic. Uncomplicated classical dengue fever was associated with 40 patients. One patient, with fatal outcome, was diagnosed as dengue haemorrhagic fever/dengue shock syndrome (DHF/DSS) grade IV, according to World Health Organization criteria (1987), as described by Nogueira et al. (1991). The New Guinea C strain, at the 5th passage, was obtained from San Juan Laboratories, CDC, Puerto Rico. Virusisolation was carried out by inoculation of human acute phase sera in monolayer cultures of C6/36 Aedes albopictus cells and identified as DEN-2 by indirect immunofluorescence assay with type specific monoclonal antibodies. Infection of cell cultures Monolayer cultures of C6/36 Ae. albopictus cells were inoculated with 50 l of each tested human positive serum, diluted 1:10 in Leibovitz-15 medium. The cultures were incubated at 28 C and fixed at the second or third day when 30-50% of cytopathic effect was observed by light microscopy. Several DEN-2 virus isolated were passed subsequently four to five times in the same cell culture line. Electron microscopy The infected cell cultures were fixed with phosphate buffered 1% glutaraldehyde, post-fixed with 2% osmium tetroxide in the same buffer, scrapped from the glass vials, washed, dehydrated in increasing acetone concentrations and included in Epon. All the steps were processed in Eppendorf centrifuge tubes. Ultrathin sections were classical stained with uranyl acetate and lead citrate and observed in a Zeiss EM-900 electron microscope. At least 50 different sectioned cells were analyzed for each sample.
TABLE 1: Distribution of DEN-2 isolates in relation to infected cells in culture, based on EM-observations of 50 cells for each isolate % of infected Number Fatal Number of isolates cells in culture of isolates with fuzzy coated by light and isolates virus like particles EM-observations 0 - 30% 12 0 10 30 - 50% 13 0 12 50 - 100% 16 1 15 Total 41 1 37 RESULTS Electron microscope observations of the Brazilian DEN-2 isolates show that classical and defective virus particles are always present at corresponding proportions of infected cells (Table); smooth membrane structures (SMS) are always more frequent than virus particles. After entering the cell by endocytosis, viral RNA was liberated into the cell cytoplasma by lowering the pH of the endocytic system and the consequent fusion of their membranes with the viral envelope, when virus replication is starting. Viral nucleic acids protrude into the RER together with the synthetised viral proteins, forming groups of typical smooth membrane enveloped virus particles inside (Fig. 1). Numerous SMS, like small vesicles or tubules, bud simultaneously also into the lumen of the RER (Fig. 2); their apparently proteinous nature was not yet defined. Another type of probable virus-like particles goes along with this process of DEN-2 virus replication. As previously described (Barth 1991), these particles present also a dense core, but are surrounded by a fuzzy structured envelope (Fig. 2). These fuzzy coated virus-like particles were initially thought to be antibodies associated to virus particles of secondary infection (Fig. 3), but they occur just in the 4th (isolates No. 39056, 40122, 39343) and the 5th (isolates No. 39357, 40247) infected cell passages executed. Detailed observations of virus replication in mosquito cells inoculated with the serum from the fatal case, reveal different aspects. More virus particles are found inside the RER compartment or RER derivated vesicles, beside the SMS. The major number of particles are coreless and present only the proteinous envelop. Fuzzy coated particles are more frequently observed than in the other DEN-2 isolates examined. Concentric disposed RER compartments, as well as an increased number of Golgi complexes (Fig. 4) are typical of this occurrence. Exocytosis is easier to be detected than in the other isolates examined. Only classical featured dengue virus particles were detected in the New Guinea C strain and a great amount of SMS fills cytoplasmic vesicles and vacuoles. DISCUSSION Ultrastructural aspects of DEN-1 virus replication inside mosquito cell cultures from the 1986 outbreak were previously described (Barth & Schatzmayr 1992) and a previous report on DEN-2 virus features was also presented (Barth 1991). Observations ofseveral isolates of DEN-2 from the Rio de Janeiro epidemic, including one fatal case, results in additional information, that complement the proposed steps (Barth 1992) of virus replication inside the cell cultures. Enhancement of virus infection by a sequential infection of different dengue virus serotypes was presented first by Halstead (1981). EM observations (Barth 1992) confirm that a major number of virus particles penetrates the cells, when DEN-2 viruses were pre-incubated with the convalescent serum from patients who have had primary DEN-1 infections, so that larger amounts of newly formed virus particles occur within the cysternae of the RER. More particles are transferred to the Golgi complex, exocytosis of virions is frequently observed and near the totality of cells in culture are infected. This means more fulminant viral expression. Antibodies uptake by the cell is limited to the first infection, while subsequently infected cell cultures, until the 5th passage, do not present any virus-antibody linked structures. So, the fuzzy coated virus-like particles cannot be related with antibodies. Figure 1: DEN-2 virus inside a mosquito cell in culture. Virus particles (V) are protruding into the rough endoplasmic reticulum (RER), 18000x. Fig. 2: numerous smooth membrane structures (SMS), typical dengue virus particles (V) and fuzzy coated particles (arrows) occur inside the RER cisternae, 40000x. Fig. 3: antibody (A)-linked dengue virus particles absorbed to the mosquito cell membrane (M), 100000x. Fig. 4: typical concentric disposed RER tubules and cisternae, linked to a golgi complex (G), 30000x (bars=200nm). SMS, like vesicles and tubules inside the RER, are probably of proteinous nature; no nucleic acid, as supposed by Ng (1987), could be confirmed inside them (Barth & Schatzmayr 1992). They appear before any new virus particle is detected in the RER and appoint the end of the latent period of virus replication. They are never released from its surrounding vesicles during cell lysis. At the moment we will consider these structures as a cell response to the viral infection. They are so common for DEN-2 as for DEN-1 infections; no alteration of number and structure of the SMS were observed at the fatal case. A great number of defective virus particles inside the RER, presenting only the viral envelopes and lacking partially or entirely its RNA, characterize the virus replication in the fatal case; the increased RER, containing a major number of virus particles and SMS, reflects the hopeless activity of the infected cells and syncytia. Increasing number of Golgi complexes means that more virus particles can be transferred from the RER to the Golgi cysternae for final glycosylation and cell exocytosis and for further cell infections. Morphology comparison of viruses structures from the 41 isolates of the Brazilian strain with the referencial New Guinea C strain reveals that the two are different. Sequencing of some of these cases, by the analysis of the E/NS1 gene junction (Rico-Hesse 1990), including the fatal one, confirmed the similarity of the Brazilian cases with the Jamaican topotype strain (genotype group III); genetic relationships separate this topotype from genotype group II, the New Guinea C reference strain. At the other hand, a relative close identity has been detected by the analysis of the E-gene fragment (Deubel et al. 1993). Our results clearly confirm a difference between the DEN-2 Brazilian virus and the New Guinea C reference strain. REFERENCES Barth OM 1991. Replication of dengue virus type 2, Brazilian strain, in mosquito cell cultures. Mem Inst Oswaldo Cruz 86: 123-124. Barth OM 1992. Replication of dengue viruses in mosquito cell cultures a model from ultrastructural observations. Mem Inst Oswaldo Cruz 87: 565-574. Barth OM & Schatzmayr HG 1992. Brazilian dengue virus type 1 replication in mosquito cell cultures. Mem Inst Oswaldo Cruz 87: 1-7. Deubel V, Nogueira RM, Drouet MT, Zeller H, Reynes JM, Ha DQ 1993. Direct sequencing of genomic cDNA fragments amplified by the polymerase chain reaction for molecular epidemiology of dengue-2 viruses. Arch Virol 129: 197-210. Halstead SP 1981. The pathogenesis of dengue. Molecular epidemiology in infectious disease. Am J Epidem 114: 632-648. Ng M 1987. Ultrastructural studies of Kunjin virus infected Aedes albopictus cells. J Gen Virol 68: 577-582. Nogueira RMR, Miagostovich MP, Lampe E, Schatzmayr HG 1990. Isolation of dengue virus type 2 in Rio de Janeiro. Mem Inst Oswaldo Cruz 85:253. Nogueira RMR, Zagner SMO, Martins ISM, Lampe E, Miagostovich MP, Schatzmayr HG 1991. Dengue haemorrhagic fever/dengue schock syndrome (DHF/DSS) caused by serotype 2 in Brazil. Mem Inst Oswaldo Cruz 86: 269. Rico-Hesse R 1990. Molecular evolution and distribution of dengue viruses type 1 and 2 in nature. Virology 174: 479-493. Schatzmayr HG, Nogueira RMR, Travassos da Rosa APA 1986. An outbreak of dengue virus at Rio de Janeiro. Mem Inst Oswaldo Cruz 81: 245-246. WHO 1987. Dengue hemorrgico: diagnstico, tratamento e controle. Organizao Mundial de Sade. Genebra. 79 p. Copyright 1994 Memorias do Instituto Oswaldo Cruz. The following images related to this document are available:Halftone images[oc94004a.gif]Photo images[oc94004a.jpg] |
| |||||||||

{kind=link}