 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 89(1): 33-40, jan./mar. 1994
Helminth Parasites of Conventionally Maintained Laboratory MiceRoberto Magalhaes Pinto+, J Julio Vicente, Dely Noronha, L Goncalves, Delir Correa Gomes+ Laboratorio de Helmintos Parasitos de Vertebrados, Departamento de Helmintologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900, Rio de Janeiro, RJ, Brasil ^+CNPq research fellows, Proc. no. 300.374/80-1 and 303.124/89-0, respectively. Received 28 June 1993, Accepted 30 November 1993
Code Number: OC94007
Sizes of Files:
Text:20K
Graphics: Line Drawings (Gif) - 155K
Photos (Jpg) 48.4K / Halftones (Gif) - 165.5K
The spectrum of intestinal parasites present in the Swiss
Webster, C57B1/6 and DBA/2 mice strains from different
animal houses was identified and prevalences compared.
Three parasites were observed during the course of this
study, namely the cestode Vampirolepis nana
(Siebold, 1852) Spasskii, 1954 (= Hymenolepis
nana) and the nematodes Aspiculuris tetraptera
(Nitzsch, 1821) Schulz, 1924 and Syphacia obvelata
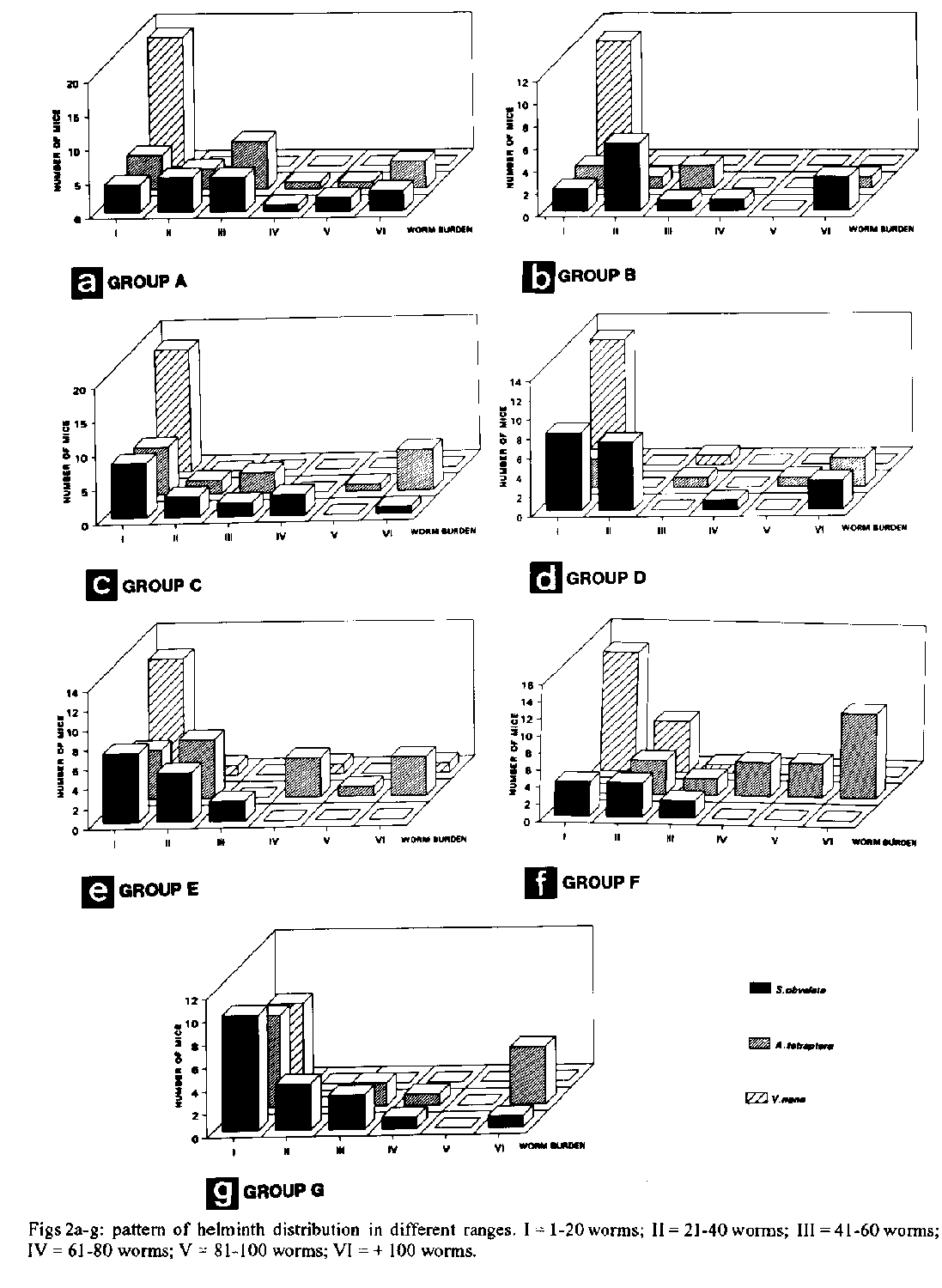
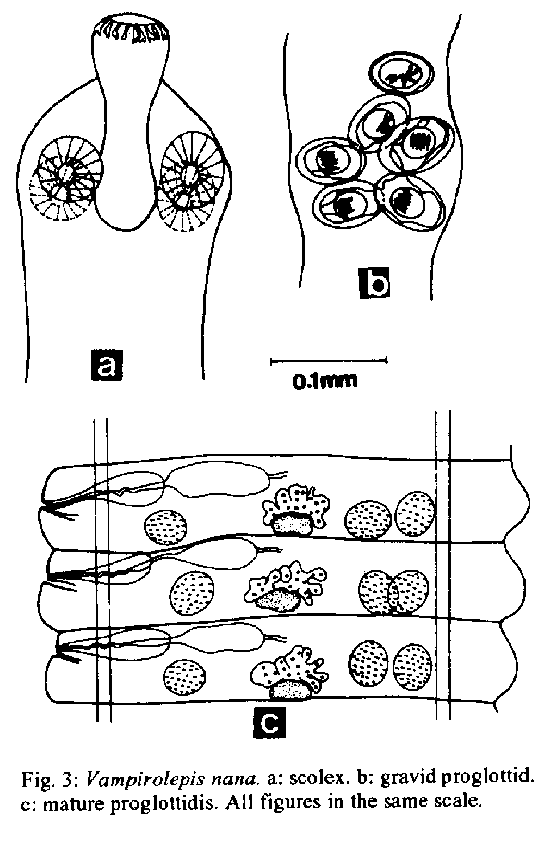
(Rudolphi, 1802) Seurat, 1916.The scope of this investigation has been widened to also include morphometric data on the parasites, to further simplify their identification, since the presence of helminths in laboratory animals is regarded as a restricting factor for the proper attainment of experimental protocols. Key words: Vampirolepis nana - Hymenolepis - Aspiculuris tetraptera - Syphacia obvelata - laboratory mice - prevalence - morphometrics - Mus musculus It is widely known that under semi-open or conventional housing, laboratory rodents are frequently infected with intestinal helminths (Eaton 1972, Hasslinger & Wiethe 1987, Cheng & Xinmei 1990). The present study reports the necropsy findings in inbred and outbred mice from institutional animal houses and was undertaken to provide data on the prevalence, distribution and morphology of the cestode Vampirolepis nana (Siebold, 1852) and the nematodes Aspiculuris tetraptera (Nitzsch, 1821) Schulz, 1924 and Syphacia obvelata (Ru- dolphi, 1802) Seurat, 1916, that are the most commonly encountered intestinal parasites in the mouse. MATERIALS AND METHODS Animals - Inbred (C57B1/6, DBA/2) and outbred (Swiss- Webster) Mus musculus L. mice strains were obtained from different institutional animal houses in Rio de Janeiro, RJ and Belo Horizonte, MG, Brazil. The suppliers were not identified by name due to ethical reasons. Twenty-five adult male host specimens from each source were necropsied for helminths and divided into seven groups, according to the supplying animal house, as follows: SW mice - groups A, B, C, D, E; C57B1/6 - group F; DBA/2 - group G. Parasites - Mice were euthanized in an ether chamber, according to the Brazilian principles for the care and use of laboratory animals (APA 1989). Helminths were recovered alive in Petri dishes, containing 0.8% NaCl solution. Nematodes were fixed with hot 10% formaldehyde solution. Cestodes were kept overnight in tap water, in the refrigerator, and transferred to cold formalin. Specimens were dehydrated in ethanol (70 GL - 100 GL), stained with carmine and cleared in phenol or beechwood creosote. Some were preserved in balsam as whole mounts and deposited in the Helminthological Collection of the Instituto Oswaldo Cruz (CHIOC). Illustrations were performed with the aid of a drawing tube connected to an Olympus light microscope. Measurements are in micrometers, unless otherwise indicated. Worm burden counting was conducted under stereoscope microscope and six ranges of distribution were considered, to include the number of worms/mouse as follows: I: 1-20 worms; II: 21-40; III: 41-60; IV: 61-80; V: 81-100; VI: + 100. Histology - Fragments of intestine were removed and immediately fixed in formalin. The material was then routinely processed for parafin embedding. Five m thick sections were stained with haematoxylin and eosin. Photomicrographies - Histological sections - obtained in an Olympus microphotographic system; helminths - obtained with a Nikon F2 camera, 55 mm microlens, connected to a 2x ring. Graphics - Harvard Graphic Program 2.3. Figs 1a-g: prevalence of helminth species. A = Aspiculuris tetraptera; S = Syphacia obvelata; V = Vampirolepis nana. Figs 2a-g: pattern of helminth distribution in different ranges. I = 1-20 worms; II = 21-40 worms; III =41-60 worms; IV = 61-80 worms; V = 81-100 worms; VI = + 100 worms. (Siebold, 1852) Spasskii, 1954 (= Hymenolepis nana) (Figs 3a-c, 6a-b, 7a-b) RESULTS The classification adopted herein is that of Petter and Quentin (1976) for the nematodes and of Schmidt (1986) for the cestode. Morphometric data are simplified and adapted. Fig. 3: Vampirolepis nana. a: scolex. b: gravid proglottid. c: mature proglottidis. All figures in the same scale. Fig 4: Aspiculuris tetraptera. a: anterior portion. b: posterior extremity of male, ventral view. c: posterior extremity of male, lateral view. d: posterior extremity of female, lateral view. e: egg. Morphometrics (based on seven specimens). Cestoda, Hymenolepididae, Hymenolepidinae. Long worms, white when alive. Body flattened, 25-80 mm long. Suckers 126 x 108. Rostelum well developed, with a circle of about 22 Y- shaped hooks, 18 long. Proglottids numerous, transversely elongated: immature 108-187 x 18-27, mature 432-561 x 52- 72 and gravid 374-504 x 144-172. Testes three, 21-32 x 25-39, arranged in transverse row or triangle. Cirrus pouch 54-72 long. Gravid uterus extending transversely. Eggs 46-54 x 36-42.
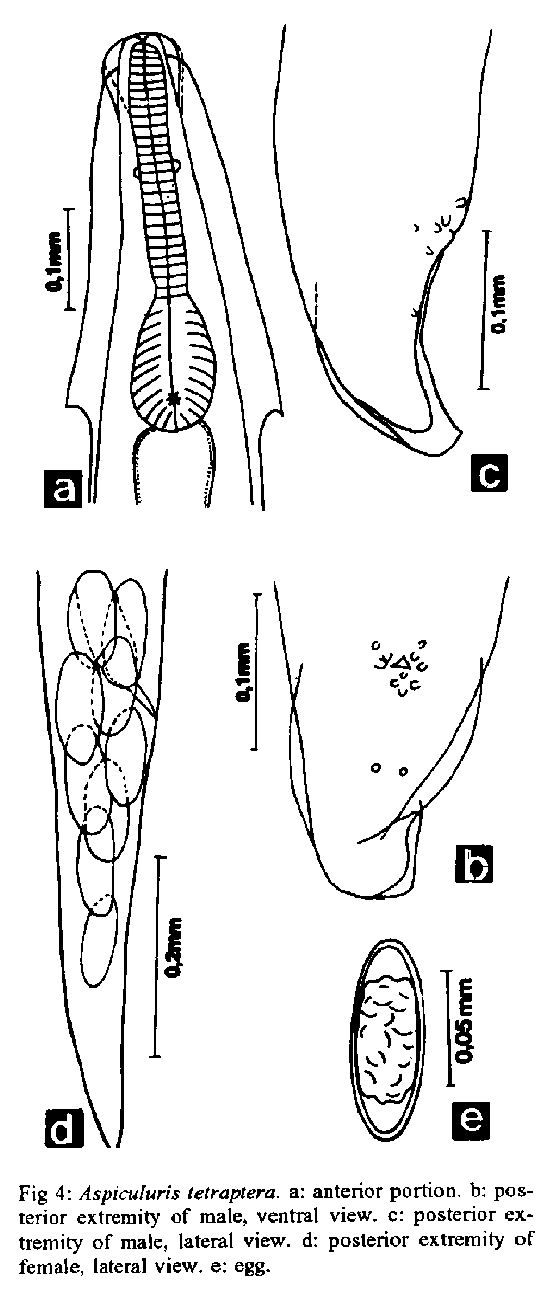
Aspiculuris tetraptera (Nitzsch, 1821) Schulz, 1924 (Figs 4a-e, 6c) Morphometrics (based on ten specimens: five males and five females). Oxyuroidea, Heteroxynematidae, Heteroxynematinae. Small worms, white when alive. Males: 2.4-3.1 mm long, 175-210 wide. Esophagus, with bulb, 288-360 long. Nerve ring and excretory pore 115-140 and 619 from anterior end, respectively. Twelve caudal papillae. Spicules and gubernaculum absent. Cloacal aperture 144-172 from posterior end. Females: 3.1-3.6 mm long, 175-245 wide. Esophagus, with bulb, 331-432 long. Nerve ring and excretory pore 158-216 and 734-936 from anterior end, respectively. Vulva 1.1-1.4 mm from anterior extremity. Eggs 72-90 x 36-53. Anus 360-486 from posterior end. Fig. 5: Syphacia obvelata. a: anterior portion. b: posterior extremity of male, ventral view. c: egg. d: posterior extremity of male, lateral view. e: posterior portion of male, lateral view. f: posterior extremity of female, lateral view.
Taxonomic summary Host: Mus musculus L. Site of infection: intestine Distribution: cosmopolitan Specimens studied: CHIOC no. 33.047 a-j (whole mounts).Syphacia obvelata (Rudolphi, 1802) Seurat, 1916 (Figs 5a-f, 6d) Morphometrics (based on ten specimens, five males and five females). Oxyuroidea, Oxyuridae. Small worms, white when alive. Great sexual dimorphism. Male: 1.0-1.1 mm long, 57-72 wide, with three "mamelons" on the ventral surface of posterior portion. Esophagus, with bulb, 144-158 long. Nerve ring and excretory pore 90-97 and 79 from anterior end, respectively. Single spicule, 72-82 long. Gubernaculum 25-39 long, with a hook-shaped distal prolongation. Three pairs of caudal papillae. Cloacal aperture 72-90 from posterior end. Females: 4.5-5.0 mm long, 245-350 wide. Esophagus, with bulb, 316-324 long. Nerve ring and excretory pore 82-90 and 86 from anterior end, respectively. Vulva 662-806 from anterior extremity. Eggs 118-126 x 43-54. Anus 518 from posterior end.
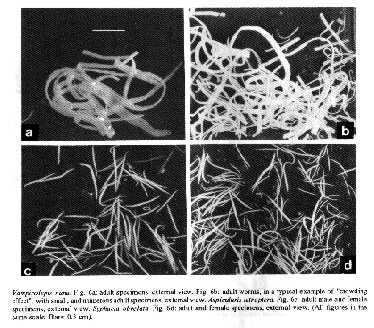
Taxonomic summary Host: Mus musculus L. Site of infection: intestine Distribution: cosmopolitan Specimens studied: CHIOC no. 33.048 a-j (whole mounts). Regardless of the source, a high degree of parasitism (96-100%) was observed in the necropsied mice. The cestode species V. nana, in single infections, appeared with the highest prevalence in group B (Fig. 1b) and associated to the nematodes A. tetraptera or S. obvelata in all groups. Specimens of V. nana were either attached to the intestinal mucosa or free in the lumen (Figs 7a-b). The highest prevalence regarding the association of V. nana and A. tetraptera was observed in the C57B1/6 mice of group F (Fig. 1f), while that of V. nana and S. obvelata occurred in group D (Fig. 1d). Fig.6:Vampirolepis nana. Fig. 6a: adult specimens, external view. Fig. 6b: adult worms, in a typical example of "crowding effect", with small, and numerous adult specimens, external view. Aspiculuris tetraptera. Fig. 6c: adult male and female specimens, external view. Syphacia obvelata. Fig. 6d: adult and female specimens, external view. (All figures in the same scale. Bars: 0.3 cm). Syphacia obvelata, in single infections, appeared with the highest prevalence in group B (Fig. 1b) and A. tetraptera in group E (Fig. 1e). The association of these nematodes with higher prevalence occurred in group G (Fig. 1g). Very high prevalences of the three species, V. nana, A. tetraptera and S. obvelata, in association, were detected in all groups, except for the group B, where it did not occur (Fig. 1b) and group D, in which a low prevalence was observed (Fig. 1d). The highest prevalence of the association was that of group A (Fig. 1a), followed by groups C (Fig. 1c), E (Fig. 1e), F (Fig. 1f) and G (Fig. 1g). In respect to the distribution of helminths, we can observe that V. nana was mainly outstanding in range I (Figs 2a-f), in all groups, except for the group G, where the three species appeared with similar patterns in this range (Fig. 2g). Aspiculuris tetraptera and S. obvelata were distributed quite regularly in all ranges in group A (Fig. 2a), while in the other groups, their distribution was not observed in some ranges (Figs 2b-g). The greatest number of infected mice with A. tetraptera, occurred in range VI in group F (Fig. 2f) and those with S. obvelata in range I in group G (Fig. 2g). DISCUSSION Experimental animal models maintained in laboratories are subject of concern, considering the role they play in the evaluation of biological parameters in the different fields of scientific research. Taking this into account and depending on the assay under development, the presence of helminths in these hosts, for example, must be regarded as a restricting factor for the proper attainment of experimental protocols. Fig. 7: Vampirolepis nana. a: cross-section of small intestine, showing worms in the lumen attached to the mucosa (arrow), 50X. b: detail of scolex (arrow) attached to the mucosa. 400X. Intestinal damage due to cestode infections in the vertebrates has been recently discussed (Hayunga 1991) and although no gross lesions were found associated with oxyuriasis in mice, it has been reported that microscopic examinations reveal the presence of immature worms in mucosal glands and increased numbers of lymphocytes and leucocytes in the colon mucosa and also that young inbred mice with heavy infections of oxyurids are thin and emaciated (Eaton 1972). Furthermore, the release of their multiple antigens in the parasitized organism, is sufficient to, temporarily, disable this individual as a suitable candidate for specific immunological assays, in which a wide range of antigenic derived cross-reactions may occur, if the presence of different groups of parasitizing agents is not avoided or previously diagnosed. It has been most usual to verify that, among the commonly used laboratory mammals from several supplying animal houses, some are heavily parasitized with helminths at the occasion of the delivery, or become infected in the laboratories of destination, where they are sometimes kept for long periods, depending on the experimental design to be accomplished. These infective processes are due to the presence of viable eggs and/or potential intermediate hosts that are not avoided as contaminants of food and wood shavings in the cages where animals are kept, in a permanent menace to the whole colony, mainly considering that the oxyurid nematodes A. tetraptera and S. obvelata have monoxenous life-cycles (Hugot 1980, 1988) whereas the hymenolepidid V. nana, with heteroxenous life- cycle, develops in the intermediate hosts, that are common insects living in mice food (Rêgo 1965, Voge & Heyneman 1957). This explains the constancy of infection with the three species, even though with low countings (range I), in all groups, except for group F, where the mice were not infected with A. tetraptera in this range, although were parasitized with this species regarding the remaining ranges (Fig. 2f). However, when groups are analyzed as a whole, the presence of A. tetraptera or S. obvelata is observed in all ranges of distribution, except for the range V in groups B, G (Figs 2b, g). Moreover, although the life-cycle of the oxiurids is monoxenous, the passage of their larval stages through the outside environment is sometimes so much reduced that it may be regarded as potentially non-existing (Hugot 1980). With basis on the results, it was verified that the present situation is critical, taking into account the patterns of helminth distribution in the examined hosts (Figs 2a-g). It was surprizing to observe the high prevalence of the three species, V. nana, A. tetraptera and S. obvelata, when they appeared associated either in the outbred (groups A, C, D, E) (Figs 1c-e) or in the inbred mice (groups F, G) (Figs 1f, g). This association was not observed only in the SW mice of group B (Fig. 1b). The "crowding effect" was detected in mice with single infections due to V. nana, with worm burdens included in ranges IV and VI of worm distribution in mice of group E (Fig. 2e). The worms, although adult and mature were very small, numerous and, at first sight, did not seem to be V. nana (Fig. 6b). This situation has been analyzed, when the effects of number and age of specimens of the cestode Hymenolepis diminuta in the nature of infections in rats are discussed (Chandler 1939). With respect to the classification of this cestode species, we based our decision on the criteria adopted by Schmidt (1986), mainly considering morphological aspects related to the presence, in the genus Vampirolepis Spasskii, 1954, of a well developed rostellum, with rostelar sheat and a circle of variable number of Y- shaped hooks. The diagnosis of the genus Hymenolepis Weinland, 1858, regarding the same characters, reports to an absent or unarmed rudimentary rostellum. With basis on these arguments, with which we agree for the moment, the species is, actually transferred from Hymenolepis and included in Vampirolepis. The main approach of this investigation is addressed to those not so much acquainted with helminthological findings during the development of experimental assays, in which laboratory animals are not supposed to harbour unexpected infections. When they occur, however, their recognition is promptly achieved, through a stereoscope microscope (Figs 6a-d). Helminth infections of laboratory animals are a common problem particularly in conventional animal facilities. They are difficult to eradicate, but if adequately controlled are considered to be relatively non pathogenic (Higgins-Opitz et al. 1990). Although most common, cosmopolitan, with high incidences and prevalences of infection, these helminths are poorly studied, mainly in Brazil, in regard to morphometric aspects. Considering this fact, the present investigation was also scheduled to provide facilities concerning the proper identification of the parasites herein reported, reinforcing the statement of Higgins-Opitz et al. (1990), when they afirm that it is very important to have the determination of the spectrum and prevalence of intestinal parasites in a rodent colony. ACKNOWLEDGEMENTS To the institutions for the supply of mice; to Dr Jose Jurberg, Department of Entomology and Dr Guido Vidal Schaeffer, Department of Helminthology, Instituto Oswaldo Cruz/FIOCRUZ for microphotographies; to Mara Lucia de Souza Lemos, IOC research fellow from the "Setor de Programacao Visual (SICT/FIOCRUZ)" for revision concerning figures and graphics herein presented. REFERENCES Apa 1989. Codigo de etica experimental com animais. SOZED, Rio de Janeiro, 8 p. Chandler AC 1939. The effects of number and age of worms on the development of primary and secondary infections with Hymenolepis diminuta in rats, and investigation into the nature of "premunition" in tapeworm infections. Am J Hyg 29: 105-114. Cheng G, Xinmei Q 1990. Observation on intestinal parasites of laboratory mice. J Shangai Agr Coll 8: 125-130. Eaton GD 1972. Intestinal helminths in inbred strains of mice Lab Animal Sci 22: 850-853. Hasslinger MA, Wiethe T 1987. Zum Oxyurenbefall kleiner Labortiere und seiner Bekampfung mit Ivermectin. Tierarztl Prax 15: 93-97. Hayunga EG 1991. Morphological adaptations of intestinal helminths. J Parasitol 77: 865-873. Higgins-Opitz SB, Dettman CD, Dingle CE, Anderson CB, Becker PJ 1990. Intestinal parasites of conventionally maintained BALB/c mice and Mastomys coucha and the effects of a concomitant schistosome infection. Lab Animals 24: 246-252. Hugot JP 1980. Sur le genre Aspiculuris Schulz, 1924 (Nematoda, Heteroxynematidae), oxyures parasites de rongeurs Muroidea. Bull Mus Natn Hist nat Paris 4th ser. A 3: 723-735. Hugot JP 1988. Les nématodes Syphaciinae, parasites de rongeurs et de lagomorphes. Taxonomie. Zoogeographie. Evolution. Mem Mus Natn Hist nat (A) 141: 1-153. Petter AJ, Quentin JC 1976. Keys to the genera of the Oxyuroidea. p. 1-30. In R C Anderson, A G Chabaud, S Willmott (eds). CIH Keys to the nematodes parasites of vertebrates. Part 4. Commonwealth Agricultural Bureaux, England. Rego AA 1965. Sobre a evolucao de Hymenolepis microstoma (Duj., 1845) em Strongylopsalis mathurinii Ribeiro, 1931, um novo hospedeiro intermedierio (Cestoda, Hymenolepididae; Dermaptera, Labiidae). Rev Brasil Biol 25: 223-231. Schmidt GD 1986. Handbook of tapeworm identification. CRC Press, Inc. Boca Raton, Florida, 675 p. Voge M, Heyneman D 1957. Development of Hymenolepis nana and Hymenolepis diminuta (Cestoda, Hymenolepididae) in the intermediate host Tribolium confusum. Univ Calif Publ Zool 59: 549-580. Copyright 1994 Memorias do Instituto Oswaldo Cruz The following images related to this document are available:Halftone images[oc94007g.gif] [oc94007f.gif]Photo images[oc94007g.jpg] [oc94007f.jpg]Line drawing images[oc94007c.gif] [oc94007a.gif] [oc94007e.gif] [oc94007b.gif] [oc94007d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}