 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 89(1 ): 63-67, jan./mar. 1994
WR Catapani/+, ER Parise, MMBL Mor, M Sadigursky*, ZA Andrade*, OA Sant'anna**, SJ Miszputen, M Siqueira** Disciplina de Gastroenterologia, Escola Paulista de Medicina, Rua Botucatu 740, 04023-900 Sao Paulo, SP, Brasil *Centro de Pesquisas Goncalo Moniz - FIOCRUZ, Salvador, BA, Brasil **Laboratorio Especial de Imunogenetica, Instituto Butantan, Sao Paulo, SP, Brasil This work was partially supported by Fundacao de Amparo a Pesquisa do Estado de Sao Paulo - FAPESP. OA Sant'Anna and M Siqueira are supported in part by Conselho Nacional de Pesquisas Cientificas e Tecnologicas - CNPq. +Corresponding author Received 28 April 1993, Accepted 28 December 1993
Code Number: OC94012
Sizes of Files:
Text: 22.4K
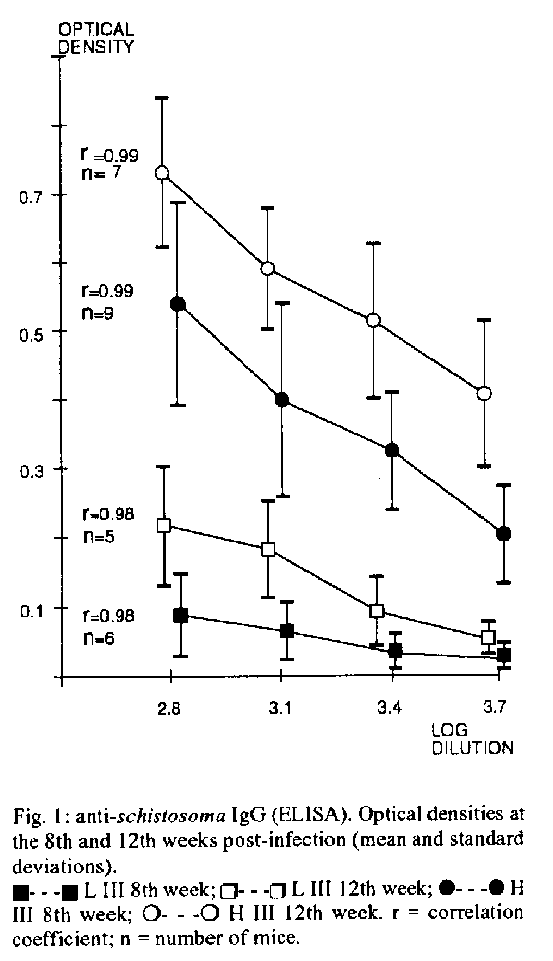
Graphics:Line Drawings (Gif) 20.4K Selection III mice have particular immunological characteristics: they are high (H 111) or low (L 111) antibody producer animals, yet both lines display similar T cell responses and macrophage activities. We submitted these mice to infection with Schistosoma mansoni 10 assess in vivo parasite and egg burden, hepatic collagen and cellular composition of granulomas in both lines. Titration of anti-Schistosoma IgG by ELISA showed remarkably higher values in H III line, at both studied periods (8th and 12th weeks post-infection). Nevertheless, the number of adult worms recovered from the portal system was similar in both lines, being not associated with anti-Schistosoma antibody levels. There is an increase in hepatic collagen from the 8th to the 12th weeks post-infection, which is paralleled by an increase in the number of eggs in the liver. This association apparently occurs at the same ratio in H III and L III animals. The most important difference found between the two lines was lhe outstanding contrast in terms of volume and eosinophil counts in the granulomas, with lesions from H III mice clearly being larger and containing more of these cells than LIII lesions. Key words: Schistosoma mansoni - collagen - granulomas - schistosomiasis - high and low antibody mice The infection by Schistosoma mansoni challenges the host with a multiplicity of antigens originated from cercariae, schistosomula, adult worms and eggs. The stimulation of the immune system by these antigens may result in a broad spectrum of responses that can be protective to the host or, conversely, potentially harmful to it. The interaction between these responses is very significant in determining the evolutionary pattern of the disease, being clear that the host's immunological characteristics are of paramount importance in this balance. A variety of murine experimental models have been used so far for in vivo studies on schistosomiasis. The present work aimed at an initial evaluation of some parasitological and biochemical parameters in a murine model bearing interesting immunological peculiarities. Starting from different outbred populations, by means of bidirectional genetic selective breeding. Siqueira et al. (1976) obtained two lines of mice characterized by high (H III) or low (L III) capacity of antibody production against flagellar Salmonella antigens. Later, it was seen that this humoral response is multispecific, that is, the same pattern of anti-SalmoneIIa antibody production is also observed with other antigenically non related immunogens (Siqueira et al. 1977). However, antibody production is the final result of complex interactions between macrophages, T cells and B cells. Hence, further research was carried out to investigate the role of these cells in the two lines. It was verified that, contrasting with the outstanding difference concerning antibody titers, phagocytosis and capacity of presenting antigens by the macrophages in H III and L III were comparable, as well as the rejection time of grafts, proliferative T-cell responses to mitogens and expression of delayed hypersensitivity type reactions (De Oliveira et al. 1985, Ferreira et al. 1985). The remarkable interline difference in terms of immunoglobulin titers besides a similar macrophage activity and T-cell responses called our attention on these mice. Considering the fundamental importance of the interaction between the parasite and host immune system, we attempted to verify in vivo the behaviour of these high or low antibody producer animals, when infected with S. mansoni, in terms of parasite burden, number of eggs in the liver, total hepatic collagen, cellular composition and volume of granulomas, assessed at two distinct evolutionary periods. MATERIALS AND METHODS Mice and infection - H III and L III female mice, 60 to 80 days old, were infected individually by tail imersion with 20 cercariae of S. mansoni, Belo Horizonte strain, recently shed from Biomphalaria glabrata snails, at the Parasitology Department laboratory, UNICAMP, Campinas, Brazil. Procedures - All the procedures were carried out at pre-determined times (8 and 12 weeks after infection), as described below. At each of these periods, respectively 9 H III and 7 L III, and 8 H III and 4 L III animals were anesthetized and bled from the retroorbital venous plexus. The mice were then sacrificed, the abdomen was opened and the portal system perfused. Adult worms were recovered and counted. Sera were used to perform individual titrations of anti-Schistosoma antibody. A second, non-bled non peffused infected batch of mice, comprising 6 H III, 5 L III and 6 H III, 6 L III mice was sacrificed respectively at the 8th and 12th week post-infection. From these animals, pieces of liver were set apart for egg counts, determination of hepatic collagen and histology. Anti-Schistosoma antibody titration - Sera from infected, ramdomly selected mice from both lines were used to perform individual ELISA tests by the usual indirect method, according to the standards of the Goncalo Moniz Research Center (FIOCRUZ), Salvador, Brazil. In short, the plates were coated with worm extract in saline (10 ug/well) and incubated overnight at 4 C. The plates were then washed and the samples placed in two-fold dilutions. After new incubation and washings, a rabbit anti-mouse IgG peroxidase conjugate (Sigma Chemical Co, USA) was applied at a 1:1000 dilution. Spectrophotometric readings were taken at 492 nm after reaction with ortho-phenylenediamine. The worm extract was prepared from male and female schistosomes homogenized in phosphate buffered saline, sonicated at 4 C for 3 min and centrifuged at 10,0000 g for 10 min. The supernatant was then collected and after protein determination, was used to coat the plates. Hepatic perfusion and recovery of worms - Performed according to Duvall and De Witt (1967). Liver egg counts and determination of collagen - The number of eggs in the liver was determined according to Cheever (1968). Only intact eggs were counted, egg remnants were not considered. Collagen was determined as described by Rojkind and Gonzales (1974). Histology - Liver specimens were fixed in 10% formalin. For each specimen, slides were done in triplicate, with a gap of 80 u between the cut levels, and stained with hematoxilin-eosin (H&E). An overall appreciation of the granulomas cellular composition was carried out on H&E stained slides. A blind approach was used throughout the histologic evaluation. Only granulomas containing a viable egg in the centre were considered, those in which the cut level was eccentric in relation to the egg were discarded. About 40-50 lesions meeting this pattern (6 to 10 per mouse) were analyzed either at the 8th week and 12th for evaluation of eosinophil numbers and measurement of granulomas, performed by using appropriate auxiliary eyepiece graticules. For eosinophil counting, only intact cells, showing the nucleus and cytoplasmic eosinophilie granules were computed. The mean number of these cells per granuloma per animal was calculated, and then the mean for each group. The volume of granulomas was calculated by measuring two diameters at right angles to each other, assuming that the egg is in the centre of the lesion. The mean granuloma volume per animal was calculated, and then the mean for the different groups. RESULTS Titration of anti-Schistosoma IgG - Fig. 1 shows the optical densities (means and standard deviation) obtained from titration of individual HIII and LIII sera, plotted against the logarithm of dilutions (dilution range 1:640 to 1:5120). It is self-evident the outstanding difference between the two lines in terms of serum anti-Schistosoma IgG levels in both periods. Recovery of worms - Table I shows total adult worms and number of pairs recovered from both lines. There is an obvious similarity between the groups, no statistically significant difference was found. Liver egg counts and hepatic collagen - Fig. 2 shows the association between the increase in hepatic collagen and egg numbers in both lines, plotting together the results from 8th and 12th weeks post-infection. The results shown indicate the increase of collagen above the mean values obtained from 4 HIII and 6 LIII control non-infected mice (respectively 1.10 and 0.98 mg collagen/gram of liver). The increase of collagen in each line was expressed by subtracting the mean normal liver collagen from the values found in the infected animals. Fig. 1: anti-schistosoma IgG (ELISA). Optical densities at the 8th and 12th weeks post-infection (mean and standard deviations). TABLE I
Recovery of adult worms by liver perfusion. Mean and standard
deviations
Worms Line 8th week 12th week
Total H III 3.44+/-1.66 (n=9) 4.25+/-2.25 (n=8)
L III 4.14+/-1.67 (n=7) 5.75+/-2.22 (n=4)
Pairs H III 1.44+/-0.72 1.50+/-0.92
L III 1.57+/-0.78 2.25+/-1.25
Analysis of variance: total worms p = 0.29; worm pairs p =
0.47; n = number of animals.
Histology - Cellular granulomas from H III mice contained
clearly more eosinophils than those from L III animals, both
at the 8th and 12th weeks. Even by simple visual observation,
these findings were distinctive enough to allow a prediction
of what line the slide being examined belonged to. Table II
shows eosinophil counts (group mean and standard deviation) in
both lines. It was also evident that central necrosis was very
frequently present in H III and almost absent in L III,
especially at the 8th week. Table III shows the granulomas
mean volumes and standard deviations for the groups in each
period studied.Fig. 2: number of eggs (x 10-1) per gram of liver versus increase in hepatic collagen (mg col lagen per gram of liver). TABLE II Number of eosinophils per granuloma. Group means and standard deviations Line 8th week 12th week H III 135.4+/-42.8 (n=6) 134.3+/-63.8 (n=6) L III 56.2+/-51.9 (n=5) 62.33+/-14.34 (n=6) Analysis of variance: p = 0.010; n = number of animals. TABLE III Volume of granulomas (mm^3 x 10^3). Group means and standard deviations Line 8th week 12th week H III 30.05+/-4.74 (n=6) 11.60+/-6.67 (n=6) L III 12.90+/-6.36 (n=5) 11.68+/-5.47 (n=6) Analysis of variance: p = 0.000; t = test for 8th week H III x L III p = 0.001; t = test for H III 8th week x 12th week; p = 0.0004; n = number of animals.DISCUSSION The main goal of this work was to correlate in vivo the parameters studied with high and low antibody production phenotypes. We expected to observe the same pattern of multi- specific response for antibody production also to S. mansoni antigens, besides those evidenced previously (Siqueira et al. 1977). This was confirmed by the ELISA test: titers of anti- Schistosoma IgG were higher in H III mice already after 8 weeks of infection, continuing to be markedly higher after 12 weeks. The large interline difference in anti-Schistosoma antibody titer could theoretically reflect on the ability of the host to kill the parasites. Previous experiments showed that, contrarily to the events in pre-infected mice, in which there is an acquired immunity that destroys a proportion of worms from a challenge infection, in naive mice a percentage of the cercariae penetrating the skin die, and no further significant loss of schistosomula or adult worms occurs thereafter, allowing a prolonged survival of parasites in the host (Sher et al. 1974, Smithers & Gammage 1980). In a single infection this situation appears to be antibody-independent, as B-cell depleted mice, artificially rendered so by treatment with anti IgM serum and infected with S. mansoni showed the same parasitic burden of their non-depleted counterparts (Cheever et al. 1985). In general lines, our data in the constitutional Selection III mice agree with the reported above. Also, considering that these animals have comparable macrophage activities (Ferreira et al. 1985) and displayed the same worm burden regardless of the anti-Schistosoma antibody titers, our results suggest a more important role played by macrophages rather than humoral immunity in determining the worm burden in a primary infection. The eggs trapped in the liver parenchima stimulate the fonnation of granulomatous lesions around them. Periovular fibrosis is the final outcome of this event. Therefore, the number of eggs in the liver play a preponderant role in determining hepatic damage. In accordance with previous data (Cheever et al. 1983), our results show an appreciable increase in the number of eggs per gram of liver, from the 8th to the 12th week, which is paralelled by an increase in hepatic collagen. The ratio (increase of collagen/number of eggs) in the liver is analogous in both groups (Fig. 2.). However, this association is expected to be only approximate, since we have counted exclusively intact eggs but not egg remnants, possibly originated from old fibrotic lesions, which actually contributed to collagen measurement. The mean total liver weight was quite similar in both lines, either at the 8th and 12th weeks (analysis of variance: p = 0.86, data not shown), therefore total collagen per liver was also comparable. Regarding the cellular composition of granulomas, the most prominent finding concerned the number of eosinophils. Although normal granulomas containing macrophages, lymphocytes and eosinophils formed in both lines, H III clearly contained more of these cells than L III lesions (Table II). H III granulomas at the 8th week are also remarkably larger than L III (p = 0.001; Table III). The number of eosinophils in the granulomas and the formation of these lesions in schistosomiasis are clearly influenced by T-dependent mechanisms. Previous investigations showed that lack of T cell functions significantly supressed the formation of granulomas, these structures being smaller and devoid of eosinophils (Hsu & Hsu 1976, Phillips et al. 1977, Mathew & Boros 1986). In Selection III mice, the CD 4+/CD 8+ ratio is similar in lymph nodes from both lines. Also, the production of interleukin 2 by CD 4+ cells under antigenic stimulation is equivalent in the two lines, or even higher in L III animals (Reis et al. 1992). It is known that T helper cells can be classified into two subsets: Th[1], producing IL-2, gamma interferon and lymphotoxin, and Th[2] that produces IL-4, IL-5 and IL-6 (Stevens et al. 1988). Considering the above information, it is tempting to speculate that in Selection III mice the differences in granuloma size and eosinophil contents at the 8th week period could be due to different subsets of T cells, with Th[2] predominating in H III animals. In accordance with our data, this hypothesis is supported by previous experiments showing that treatment of mice with anti-IL-5 monoclonal antibody did not affect worm burden, counting of eggs in the tissues and hepatic fibrosis, but reduced dramatically the number of eosinophils in the 8th week granulomas, and the volume of these lesions (Sher et al. 1990). It was also evident that modulation of granuloma size ocurred efficiently in H III line, with a reduction of about 38.6 % of volume from the 8th to the 12th week (p= 0.0004, Table III), but did not occur at all in the L III line. Again this effect could be influenced by different citokines produced in the granulomas in each line. However, the presence of central necrosis in the granulomas of H III animals may suggest the formation of local immune complexes, indicating that the participation of antibodies in the modulation process cannot be ruled out. In conclusion, Selection III mice seemed to provide an interesting model for further studies on experimental schistosomiasis, specially concerning the immunopathology of granulomas. Among the parameters studied in these mice with a single S. mansoni infection, the most remarkable peculiarity between high or low antibody producer animals was the distinctive pattern in terms of volume and eosinophil contents in these lesions. There is also a more marked necrosis in the initial H III granulomas, less evident in L III animals. Worm burden was not related with capacity of antibody production, and increase in liver collagen seemed to occur at the same rate in H III and L III lines. ACKNOWLEDGMENTS To Dr Ana Maria Guaraldo (Unicamp) for infecting the mice, and to Mr John Bode (GI Labs, Western General Hospital, Edinburgh University, UK) for his technical help. REFERENCES Biozzi G, Mouton D, Sant'Anna OA, Passos HC, Gennari OM, Reis MH, Ferreira VCA, Heumann AM, Bouthillier Y, Ibanez OM, Stiffel C, Siqueira M 1979. Genetics of immunoresponsiveness to natural antigens in the mouse. Curr Top Microbiol Immunol 85: 31-98. Cheever AW 1968. Conditions affecting the accuracy of potassium hydroxide digestion techniques for counting Schistosoma mansoni eggs in the tissue. Bull World Health Organiz 39: 328-331. Cheever AW, Byram JE, Hieny S, Von Lichtenber F, Lunde MN, Sher A 1985. Immunopathology of Schistosoma japonicum and S. mansoni infection in B cell depleted mice. Parasite Immunol 7: 399-413. Cheever AW, Dunn MA, Dean DA, Duvall RD 1983. Differences in hepatic fibrosis in ICR, C3H, and C57B1/6 mice infected with Schistosoma mansoni. Trans R Soc Trop Med Hyg 32: 1364-1369 De Oliveira S, Ibanez O, Mouton D, Sant'Anna OA, Siqueira M, Biozzi G 1985. Independent polygenic regulation of quantitative antibody responsiveness and expression of delayed- type hypersensitivity (DTH). Clin Exp Immunogenet 2: 223-233. Duvall RH, DeWitt WB 1967. An improved perfusion technique for recovering adult schistosomes from laboratory animals. Ant J Trop Med Hyg 16: 483-486. Ferreira V, Gennari M, Reis MH, Siqueira M, Stiffel C, Mouton D, Biozzi G 1985. Potentialities of immunocompetent cells in high and low antibody producing lines of mice obtained by selective breedings for responsiveness to flagellar or somatic antigens of Salmonellae. J Immunogenet 12: 309-319. Hsu CK, Hsu SH 1976. Immunopathology of schistosomiasis in athymic mice. Nature 262: 397-398. Mathew RC, Boros DL 1986. Anti- L3T4 antibody treatment supresses hepatic granuloma formation and abrogates antigen-induced interleukin-2 production in Schistosoma mansoni infection. lnfec Immun 54: 820-826. Phillips SM, DiConza JJ, Gold JA, Reid WA 1977. Schistosomiasis in the congenitally athymic (nude) mouse. I-Thymic dependency of eosinophilia, granuloma formation and host morbidity. J Immunol 118: 594-599. Reis MH, Ibanez OM, Cabrera WH, Ribeiro OG, Mouton D, Siqueira M, Couderc J 1992. T-helper functions in lines of mice selected for high or low antibody production (Selection III): modulation by anti-CD4+ monoclonal antibody. Immunology 75: 80-85. Rojkind M, Gonzales E 1974. An improved method for determining specific radioactivities of proline 14 C and collagen 14 C in collagen and in noncollagenous proteins. Anal Biochem 57: 1-7. Sher A, Coffman R.L, Hieny S, Scott P, Cheever AW 1990. Interleukin 5 is required for the blood and tissue eosinophilia but not granuloma formation induced by infection with Schistosoma mansoni. Proc Natl Acad Sci USA 87: 61-65. Sher A, Mackenzie P, Smithers SR 1974. Decreased recovery of invading parasites from the lungs as a parameter of acquired immunity to schistosomiasis in the mouse. J Infect Dis 130: 626-633. Siqueira M, Bandieri A, Reis MS, Sant'Anna OA, Biozzi G 1976. Selective breeding of mice for antibody responsiveness to flagellar and somatic antigens of Salmonellae. Eur J lmmunol 6: 241-249. Siqueira M, Esteves MB, Ibanez OM, Ferreira VCA, Sant'Anna OA, Reis MH, Biozzi G 1977. Nonspecific genetic regulation of antibody responsiveness in the mouse. Eur J Immunol 7: 195-203. Smithers SR, Gammage K 1980. Recovery of Schistosoma mansoni from the skin, lungs and hepatic portal system of naive mice and mice previously exposed to S. mansoni: evidence for two phases of parasite attrition in immune mice. Parasitology 80: 289-300. Stevens TL, Bossie A, Sanders VM, Femandez-Botran R, Coffman RL, Mossmann TR, Vitetta ES 1988. Regulation of antibody isotype secretion by subsets of antigen-specific helper T cells. Nature 334: 255-261. Copyright 1994 Memorias do Instituto Oswaldo Cruz. The following images related to this document are available:Line drawing images[oc94012b.gif] [oc94012a.gif] |
| |||||||||

{kind=link}
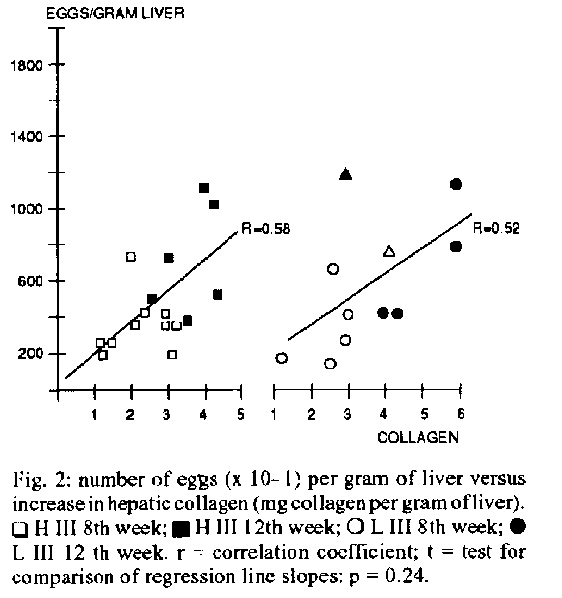{kind=link}