 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
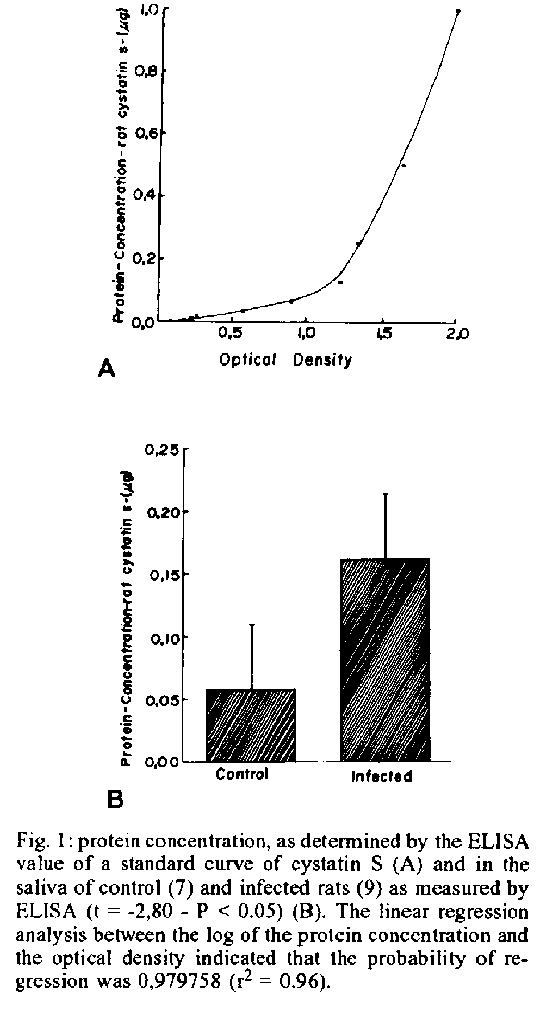
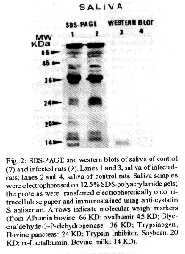
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 89(1): 81-85, jan./mar. 1994 Induction of Synthesis of the Rat Cystatin S Protein by the Submandibular Gland During the Acute Phase of Experimental Chagas DiseaseJB Alves, MSD Alves, Y Naito* Departamento de Morfologia, ICB/UFMG, Caixa Postal 2486, 31270-901 Belo Horizonte, MG, Brasil *Dept of Biological Chemistry, Faculty of Pharmaceutical Sciences, Nagoya City University, Nagoya, Japan This work was supported by grants from CNPq (040.1968/88) and PRPq/UFMG (23072.031533-70). Received 12 July 1993, Accepted 14 January 1994
Code Number: OC94014
Sizes of Files:
Text: 20.2K
Graphics:Line Drawing (Gif) 15K
Photo (Jpg) - 10K / Halftones (Gif) - 24K Rats experimentally infected with Trypanosoma cruzi Y strain exhibited hypertrophy of the submandibular gland at 18 days after infection. SDS-PAGE of infected rats saliva revealed the presence of an additional band with an apparent molecular weight of about 13KDa. Electrophoresis of protein saliva and immunochemical analysis with antibody against rat cystatin S confirmed that the protein was identical to that induced by beta adrenergic stimulation. Key words: submandibular gland - Chagas disease - rat cystatin S The observations that in Chagas disease, there occurs enlargement of the submandibular gland in rats at 18 and 32 days after infection with Trypanosoma cruzi was first reported by Alves (1977). At histologic level, the main changes observed were acinar development and delayed ductal maturation that are more evident in the developing granular ducts (Alves & Machado 1980). The enlargement induced by T. cruzi infection (acute phase) is mediated through beta-receptors since it is blocked by propranolol (Alves & Machado 1986). The histological changes observed in submandibular glands during the acute phase of Chagas disease are similar to those caused by administration of sympathomimetic drugs especially isoproterenol (Schneyer & Shackleford 1963, Ekfors et al. 1972, Yamashina & Barka 1972). Ultrastructural observations of the enlarged glands under isoproterenol (IPR) treatment revealed that the content of acinar secretory granules was altered (Takahana & Barka 1967, Kanda et al. 1968) suggesting that the secretory products of these glands may be chemically different (Robinovitchi et al. 1977). IPR not only stimulates overall protein synthesis in the submandibular gland, but also induces the synthesis of specific proteins that are absent or occur in very low concentration in unstimulated glands. In fact, a specific protein (rat cystatin S) has been identified in the submandibular gland of IPR treated-rats (Menaker et al. 1974, Naito 1981, Barka et al. 1986, Shaw et al. 1988, Shaw & Barka 1989, Shaw et al. 1990), after teeth amputation (Abe et al. 1979, Yagil & Barka 1986) and after papain administration (Naito et at. 1992). Measurements of the inhibitory activity of purified rat cystatin S revealed that this protein is a potent inhibitor of cysteine proteinases (Shaw et al. 1988). Recently, Alves (1990) reported in the acinar cells of 18 days infected-rats, secretory granules with an unusual fine structure, similar to that described by Kanda et al. (1968) in IPR treated rats. Such finding has suggested that those granules could represent secretion of rat cystatin S (inhibitor of cysteine proteinases) or another protein similar to those described under beta-adrenergic stimulation. In the present paper, biochemical methods were used in order to identify an inhibitor of cysteine proteinases as an effort to understand the role of this inhibitor in the rat submandibular gland during the acute phase of Chagas disease. MATERIALS AND METHODS Animals inoculation - Sixteen male Holtzman rats aged 27-29 days were inoculated intraperitoneally with 0.15 ml of blood containing about 300.000 trypomastigotes of Y strain of T. cruzi (Silva & Nussenzweig 1953). Living trypomastigotes were detected in the blood stream of all innoculated animals on the day 10 after infection. Saliva collection - 18 days after inoculation, the infected rats (9) and no infected controls (7) were anesthetized with an intraperitoneal injection of chloral hydrate (400 mg/kg), placed in supine position and tracheotomized. After dissecting out, the sublingual glands and ligating its duct, the left submandibular duct was cannulated with a glass cannule. Stimulation of secretion was achieved by an intraperitoneal injection of isoproterenol (16 mg/kg). Saliva was collected during 1 hr into Ependoff tubes kept on ice. Total protein was determined according to Bradford (1976). Histological study - Before stimulation, the right submandibular gland (SBM) was removed and fixed during 12 hr at 4 C with paraformaldehyde buffered in 0,1M PBS at pH 7.4. It was then dehydrated through a graded series of ethanol, embedded in glycol methacrylate JB-4 (Polysciences); sectioned at 3 um and stained with 1% toluidine blue dissolved in 1% sodium tetraborate or hematoxilin and eosin. Quantification of rat cystatin S - The concentration of rat cystatin S of SBM saliva was measured by a solid phase enzyme-linked immunoabsorvent assay (ELISA) by using a specific antibody against this protein. This antibody was raised in rabbit against rat cystatin S from saliva of IPR treated rats (Naito 1981). Briefly, wells of multi-well plates were coated with saliva. The wells were washed and the remaining sites were blocked with 1% bovine serum albumin. The incubation of the wells with diluted rabbit anti-rat cystatin S at 1:800, dilution in 1% BSA, was followed by repeated washings and incubation with peroxidase-coupled goat anti-rabbit IgG in 1:2000 dilution (Sigma). After washings, peroxidase activity was measured by the absorption at 492 nm using H2O2/ O-phenylenediamine dihydrochloride as substrate. In the ELISA described above the antigen (rat cystatin S) was quantified under conditions in which the logarithm of the concentration of the protein was linearly related to the optical density (OD). A linear regression analysis of the log protein concentration versus OD 492 indicated that probability of regression was 0.979758 (r2 = 0.96). Statistical analysis of the data was performed by means of a t test. SDS-PAGE of saliva and western blot Saliva samples of each control and infected rats were used for electrophoresis and western blot. Electrophoresis was performed in 12.5% polyacrylamide gels, according to Laemmli (1970). The electrophoretic transfer of proteins from sodium dodecyl sulfate (SDS)-polyacrylamide gels to nitrocelulose papers was carried out by using the conditions described by Burnetter (1981). The western blot were immunostained essentially as described by Barka et al. (1986). RESULTS Histological observations - The histological aspect of the normal SBM as seen in the control rats, has already been described (Jacoby & Leeson 1959, Alves & Machado 1980). The submandibular glands of the infected rats exhibit a histological picture that confirms our early finding (Alves & Machado 1980, 1986): developed acini, as judged by their size, and scanty cytoplasmic basophilia. As the adult appearance of the acini was reached earlier in the infected animals suggesting an acceleration of the acinar development. ELISA - By ELISA we have measured the relative concentration of the rat cystatin S in the submandibular gland saliva of control and infected rats. The results are shown in Figs 1A and B. There was a significant difference between the control and infected rats (Fig. lB). This results indicate that rat cystatin S is synthesized and secreted in submandibular gland. SDS-PAGE and Western blot - Cystatin S was seen as a single band with an apparent molecular weight of about 13 KD in the silverstained SDS-polyacrylamide gels of the infected rats saliva. Fig. 1: protein concentration, as detennined by the ELISA value of a standard curve of cystatin S (A) and in the saliva of control (7) and infected rats (9) as measured by ELISA (t= -2,80 - P < 0.05) (B). The linear regression analysis between the log of the protein concentration and the optical density indicated that the probability of regression was 0,979758 (r^2 = 0.96). Fig. 2: SDS-PAGE and western blots of saliva of control (7) and infected rats (9). Lanes I and 3, saliva of infected rats; lanes 2 and 4, saliva of control rats. Saliva samples were electrophoresed on 12.5% SDS-polyacrylamide gels; the proteins were transferred electrophoretically onto nitrocellulose paper and immunostained using anti-cystatin S antiserum. Arrows indicate molecular weight markers (from Albumin bovine: 66 KD; ovalbumin 45 KD; Glyceraldehyde-3-P-dehydrogenase: 36 KD; Trypsinogen, Bovine pancreas: 24 KD; Trypsin Inhibitor, Soybean: 20 KD; alpha-Lactalbumin, Bovine milk: 14 KD). Immunoblot using anti-rat cystatin S antiserum revealed a single protein band with the same electrophoretic mobility in the saliva obtained from infected rats. No trace of this protein was observed in the control saliva. SDS-PAGE and immunoblots of such gels corroborated the findings obtained by the ELISA. DISCUSSION Acinar hypertrophy accompanied by ductal atrophy, as observed in the submandibular gland of rats affected by T. cruzi (Alves & Machado 1980) is similar to those observed in glands enlarged after treatment with adrenergic agents, especially isoproterenol (Schneyer & Shakleford 1963, Ekfors et al. 1972, Yamashina & Barka 1973) or teeth amputation (Yagil & Barka 1986). The hypertrophic enlargement of the submandibular glands produced by chronic administration of pharmacological doses of IPR is also accompanied by the expression of a number of proteins and glycoproteins that are absent or occur in very low concentrations in unstimulated glands. The best characterized substances are those of the family of proline-rich proteins (Mirels et al. 1987) and a 13 KD protein (rat cystatin S) inducible by IPR injection (Naito 1981, Naito & Suzuki 1981) or teeth amputation (Yagil & Barka 1986, Abe et al. 1979) or papam administration (Naito et al. 1992). It has been suggested that in the acute phase of Chagas disease increased levels of plasma catecholamines could accelerate acinar maturation or stimulate protein synthesis (Alves & Machado 1986). Increase of the circulating levels of catecholamines would then be expected as a result of large scale destruction of adrenergic nerve terminals in the acute phase, as does occur in the heart (Machado et al. 1984) and in the parotid gland (Alves 1990). It appears, therefore, that the circulating catecholamines would act as a beta-adrenergic agonist directly on the submandibular acina cells and, like IPR, could regulate the expression of gene coding for specific proteins (Ziemer et al. 1982). Our data obtained by ELISA, SDS-PAGE and western blots are indicative of the presence in the rat submandibular gland with Chagas disease of a protein which is eletrophoretically and immunologically identical with rat cystatin S induced by IPR (Naito 1981). Although no quantitative study has been performed in order to compare levels of rat cystatin S in Chagas disease, IPR and teeth amputation, morphological data suggest that in infected rats the process was less effective than in the others. Also, as the changes produced by IPR and teeth amputation are reversible, the same happens in Chagas disease, since the enlargements returns to normal, as shown by Alves and Machado (1980). The role played by rat cystatin S and its relationship to the alterations of the submandibular gland during the acute phase of Chagas disease are yet unknown. However, increases in cystatin S concentration within tissue have been associated with pathophysiological changes, including inflammation (Brzin et al. 1984, Hopsu-Havu et al. 1983), myelin destruction (Kopitar et al. 1983), suggesting that the production of cystatin S may be a response to tissue injuries. In fact, it has been reported in rat submandibular gland, induction of cystatin S by a variety of systemic agents that are capable of causing tissue damage by a different mechanism (Cohen et al. 1993). Furthermore, in rats with Heymann nephrities, a chronic autoimmune disease cystatin was expressed within the cells of renal proximal tubules (Cohen et al. 1990). On the other hand, during acute phase of Chagas disease in spite of the absence of appreciable inflammatory process in the submandibular gland (Alves & Machado 1980), these glands undergo severe reduction of sympathetic (Machado et al. 1984) and parasympathetic innervation (Alves & Machado 1986). Together with our previous findings the present data indicate that the submandibular glands respond to denervation caused by Chagas disease in a manner similar to that observed in isoproterenol treated rats. Thus, cystatin S production may represent one possible physiological response to tissue injury. Our findings also support the hypothesis of Choen et al. (1993), in which detection of inducible cystatin may serve as a sensitive biological marker of tissue damage in salivary glands. Further studies are needed to clarify the significance of cystatin S induction in the pathogeneses of salivary glands during acute phase of Chagas disease. REFERENCES Abe K, Fujita R, Yoneda M, Yokata Y, Dawes C 1979. Changes in protein secretion by rat submandibular salivary glands after enlargements caused by repeated amputation of the lower teeth. Archs Oral Biol 24: 515-519. Alves JI] 1977. Alteracoes da inervacao autonoma e da histologia da glandula submandibular na doenca de Chagas experimental em ratos. Rev Bras Pesq Med Biol 10: 210 abstract. Alves JB 1990. Adrenergic innervation of the rat parotid gland in experimental Chagas infection. Rev Bras Cien Morfol 7: 20-23. Alves JB, Machado CRS 1980. Histologic and histoquantitative study of the rat submandibular gland in Chagas disease. Archs Oral Biol 25: 437-443. Alves JB, Machado CRS 1986. Effect of adrenoceptor blockers of the acinar enlargement induced by Chagas disease in the rat submandibular glands. Brazilian J Med Biol Res 19: 69-73. Barka T, Yagil CH, van der Noen, Naito Y 1986. Induction of the synthesis of a specific protein in rat submandibular gland by isoproterenol. Lab Invest 54: 165-171. Bradford MM 1976. A rapid an sensitive method for the quantification of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254. Brzin J, Popovic T, Turk V, Borehart U, Machleidt W 1984. Human cystatin, a new protein inhibitor of cysteine proteinases. Biochem Biophys Acta 964: 329-339. Burnette WW 1981. Western blotting electrophoresis transfer of proteins from sodium dodecyl sulfate-polyadrylamide gels to unmodified nitrocellulose and radiografic detection with antibody and radioiodinated protein A. Anal Biochem 112: 195-203. Cohen RE, Bedi GS, Neiders ME 1990. Tissue distribution of an inducible cystatin in isoproterenol-treated rats. Lab Invest 62: 452-458. Cohen RE, Neiders ME, Bedi GS, Comeau R 1993. Induction of Type 2 cystatin in rat submandibular glands by systematically administred agents. Archs Oral Biol 38: 319-325. Ekfors T, Chang WWL, Bressler RS, Barka T 1972. Isoproterenol accelerates the postnatal differentiation of rat submandibular gland. Devl Biol 29: 38-47. Hopsu-Havu VK, Joronen I, Jarvinen M, Rinne A 1982. A new cysteine proteinase inhibitor in human serum. Eur Rev Med Pharm Sci 4: 389-394. Jacoby F, Leeson CR 1959. The postnatal development of the rat submaxillary gland. J Anat 93: 201-216. Kanda T, Mayfield ED Jr, Ghidoni JJ 1968. Ultrastructural alterations in submaxillary acinar eelIs following isoproterenol administration: a new form of secretion granule. Exp Med Path 9: 189-196. Kopitar L, Stern F, Marks N 1983. Cerebrocystatin suppresses degradation of mylin basic protein by purified brain cysteine proteinase. Biochem Biophys Res Commun 112: 1000-1006. Laemmli UK 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685. Machado ABM, Machado CRS, Gomes CB 1975. Depletion of heart norepinephrine in experimental acute myocarditis caused by Trypanosoma cruzi. Experientia 31: 1202-1203. Machado CRS, Alves JB, Machado ABM 1984. Adrenergic innervation and noradrenaline content of the rat submandibular gland during the experimental Trypanosoma cruzi infection. J Neural Transmi 59: 289-297. Machado CRS, Machado ABM, Chiari CA 1978. Recovery from norepinephrine depletion in experimental Chagas disease. Am J Trop Med Hyg 27: 20-24. Menaker L, Sheetz JH, Cobb CM, Navia JM 1974. Gel electrophoresis of whole saliva and associated histologic changes in submandibular glands of isoproterenol-treated rats. Lab Invest 30: 334-341. Mirels L, Bedi GS, Dickinson DP, Gross KW, Tabak LA 1987. Molecular characterization of glutamine acid/ glutamic-rich secretory proteins from rat submandibular glands. J Biol Chem 262: 7289. Naito Y 1981. Studies on LM protein appearing in submandibular glands of isoproterenol-treated rats. I. Purification and characterization of its appearance. Chem Pharm Bull 29: 1365-1372. Naito Y, Suzuki I 1981. Studies on LM protein appearing in submandibular glands of isoproterenol-treated rats. II. Physicochemical properties. Chem Pharm Bull 29: 1373-1376. Naito Y, Suzuki I, Hasegawa S 1992. Induction of cystatin S in rat submandibular glands by papain. Comp Biochem Physiol 102B: 861-865. Robnovitchi MR, Keller P J, Johnson DA, Iversen JM, Kauffman DL 1977. Changes in rat parotid salivary proteins induced by chronic isoproterenol administration..J Dent Res 56: 290-303. Scheneyer CA, Shackleford JM 1963. Accelerated development of salivary glands of early post-natal following isoproterenol. Proc Soc Exp Biol med 112: 320-324. Shaw PA, Cow JL, Barka T, Naito Y 1988. Cloning and sequencing of cDNA encoding a rat salivary cysteine proteinase inhibitor inducible by B-adrenergic agonist. J Biol Chem 263, 18133-18141. Shaw PA, Barka T 1989. B-adrenergic induction of a cysteine-proteinases-inhibitor mRNA in rat salivary glands. Biochem J 257: 658-689. Shaw PA, Barka T, Woodin A, Shachter BS, Cow JL 1990. Expression and induction by beta-adrenergic agonists of the cystatin S gene in submandibular glands of developing rats. Biochem .J 265: 115-120. Silva LHP, Nussenweig V 1953. Sobre uma cepa de Trypanosoma cruzi altamente virulenta para o camundongo brando. Folia Cl Biol 20: 191-208. Takahana M, Barka T 1967. Electron microscopic alterations of submaxillary gland produced by isoproterenol. J Ultrastr Res 17: 452-474. Yagil C, Barka T 1986. Induction of a specific (LM) Protein in the submandibular gland of the rat by repeated amputation of the lower incisor teeth. Amer J Anat 177: 513-518. Yamashina S, Barka T 1972. Localization of peroxidase activity in the developing submandibular gland of normal and isoproterenol-treated rats. Histochem Cytochem 20: 855-872. Ziemer MA, Mason A, Carlson DM 1982. Cell-free translation of proline rich mRNA's. J Biol Chem 257: 11176-11180. Copyright 1994 Memorias do Instituto Oswaldo Cruz.
The following images related to this document are available:Halftone images[oc94014b.gif]Photo images[oc94014b.jpg]Line drawing images[oc94014a.gif] |
| |||||||||

{kind=link}
{kind=link}