 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Visceral Leishmaniasis in Teresina, State of Piaui, Brazil: Preliminary Observations on the Detection and Transmissibility of Canine and Sandfly InfectionsJA Vexenat, JA Fonseca de Castro (*), R Cavalcante (*), JP Tavares (**), MRB da Silva (*) WH Batista (***), JH Furtado Campos (*), MK Howard (****), I Frame (****), R McNerney (****), S Wilson (****), MA Miles (****/+)
Code Number: OC94028
Size of Files:
Text:21.7K
Graphics: Photographs (Jpg) - 62K / Halftones (Gif) - 88K
Universidade de Brasilia, 70910-900 Brasilia, DF, Brasil * Universidade Federal do Piaui, Campus do Ininga, Teresina, PI, Brasil ** Fundacao Nacional de Saude, Teresina, PI, Brasil *** Universidade Estadual do Piaui, Rua Joao Cabral S/No - 64002-150 Teresina, PI, Brasil **** Departments of Medical Parasitology and Clinical Sciences, London School of Hygiene and Tropical Medicine, Keppel Street, London WC1E 7HT, UK
A Leishmania donovani-complex specific DNA probe was used to confirm the widespread dissemination of amastigotes in apparently normal skin of dogs with canine visceral leishmaniasis. When Lutzomyia longipalpis were fed on abnormal skin of five naturally infected dogs 57 of 163 (35%) flies became infected: four of 65 flies (6%) became infected when fed on apparently normal skin. The bite of a single sandfly that had fed seven days previously on a naturally infected dog transmitted the infection to a young dog from a non-endemic area. Within 22 days a lesion had developed at the site of the infective bite (inner ear): 98 days after infection organisms had not disseminated throughout the skin, bone marrow, spleen or liver and the animal was still serologically negative by indirect immunofluorescence and dot-enzyme-linked immunosorbent assay. When fed Lu. longipalpis were captured from a kennel with a sick dog known to be infected, 33 out of 49 (67%) of flies contained promastigotes. In contrast only two infections were detected among more than 200 sandflies captured in houses. These observations confirm the ease of transmissibility of L. chagasi from dog to sandfly to dog in Teresina. It is likely that canine VL is the major source of human VL by the transmission route dog-sandfly-human. The Lmet2 DNA probe was a useful epidemiological tool for detecting L. chagasi in sandflies.
Key words: canine visceral leishmaniasis - Lutzomyia longipalpis - xenodiagnosis - experimental transmission - DNA probe - chemiluninescence
Human visceral leishmaniasis (VL) due to Leishmania chagasi is endemic to the semi-arid northeastern region of Brazil: canine VL is also widespread and infection rates of more than 20% have been recorded in dogs. The anthropophilic sandfly vector is Lutzomyia longipalpis. VL was first reported from State of Piaui in 1934 but in the 1980s there was a large outbreak in the State capital, Teresina, and the incidence of human VL has begun to rise again in 1992/93 (Costa et al. 1990, Tavares et al. 1993)
Traditionally the detection of L. chagasi in infected dogs and sandflies has depended on parasitological methods with the aid of serology to identify those dogs most likely to be infected. In the long term DNA probe technology is likely to provide additional tools for the rapid identification of infectious agents in large numbers of samples. The introduction of sensitive chemiluminescent detection procedures as an alternative to radiolabelling means that DNA probes will become more widely available (Wilson et al. 1992). An L. donovani complex- specific probe (Lmet2; Howard et al. 1991) has been developed as a tool to help identify the L. donovani-complex in sandflies and in samples derived from mammalian hosts (Howard et al. 1992, Gebre-Michael et al. 1993). This probe has been used successfully in the laboratory with chemiluminescence (Wilson et al. 1992) and this has suggested the production of DNA probe kits for Leishmania (McNerney et al. 1994.).
The skin is a major site of L. chagasi infection in dogs that have overt symptomatic VL and yet infection rates in wild-caught sandflies are generally reported to be rather low. The dog is considered to be the major reservoir of human disease but the presence of an infected dog is apparently not necessarily associated with the occurrence of human VL in a family: this has led to the suggestion that human-sandfly- human transmission may be an important source of human infections, although there are other explanations of the observed distribution of human VL (Evans et al. 1992, Tavares et al. 1993).
Here, we describe preliminary work with the Lmet2 DNA probe on the distribution of L. chagasi infection in naturally infected dogs, on the transmissibility of such infections and on the detection of L. chagasi in infected sandflies.
MATERIALS AND METHODS
Dogs suspected of infection with L. chagasi were either referred to us following a positive indirect immunofluorescent antibody test (IFAT) or were strays that would have been destroyed. Parasitological examination was by microscopy of Giemsa-stained skin impression smears and sternal bone marrow aspirates using standard methods. L. chagasi isolates were cultured on Difco blood-agar medium (Miles et al. 1981). Colony-bred Lu. longipalpis were used for xenodiagnoses. Peridomestic or domestic Lu. longipalpis were caught in the suburbs of Teresina either by hand using suction or battery operated 'pooters' or in CDC light traps. Xenodiagnosis flies or wild-caught flies were dissected and examined microscopically.
The Lmet2 DNA probe, either radiolabelled or with chemiluminescent detection, was used to screen tissue samples from dogs and to examine sandflies for the presence of L. chagasi. The basis of this method is described by Wilson et al. (1992) and full details of protocols are given elsewhere (Howard et al. 1991, McNerney et al. 1994). Briefly, skin biopsies from dogs were preblotted onto filter paper to remove excessive blood and then touch-blotted onto a nylon membrane (Hybond N, Amersham). Dissected sandflies were transferred to nylon membranes or undissected flies were squash-blotted directly on the nylon membrane by rolling a plastic or presoaked wooden tooth pick onto the abdomen of the fly (Gebre-Michael et al. 1993). Radiolabelled probe was produced as described by Howard et al. (1991, 1992) and digoxygenin (Boehringer Mannheim) labelled probe was produced by polymerase chain reaction (PCR) (Wilson et al. 1992).
RESULTS AND DISCUSSION
L. chagasi in symptomatic and asymptomatic dogs - Giemsa-staining of skin biopsy impression smears revealed abundant amastigotes in abnormal skin of 60% of dogs (55 of 92) with symptomatic visceral leishmaniasis. Intensive microscopy of impression smears from apparently unchanged areas of skin could detect amastigotes in some samples. Amastigotes could also be found in skin of more than 30% of dogs (18 of 57) that were IFAT positive but apparently healthy, for example, by microscopy of skin biopsies taken from the slightly raised edge of the external ear. Around 40% of symptomatic dogs (38 of 92) had parasitologically positive sternal bone marrow aspirates by microscopy of single samples (Vexenat et al. unpublished).
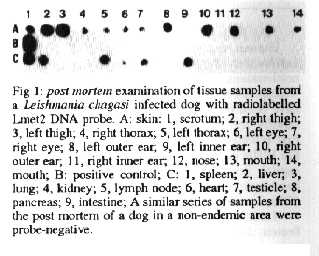
The radiolabelled Lmet2 DNA probe (Fig. 1) and Lmet2 with chemiluminescent detection (not shown, McNerney et al. 1994) were able to detect amastigotes in the skin of infected dogs, although we have not yet systematically compared probe sensitivity with parasitological and serological means of diagnosis. The probe also confirmed the widespread dissemination of amastigotes in the skin, even in skin that was apparently normal: post mortem tissue samples of one animal were probe positive with skin from the right and left outer ears (but not the inner ears), the nose, adjacent to the mouth and eyes, the right and left thorax, the right and left thigh and the scrotum; positive internal organs were the spleen, liver, lymph node, intestine, heart and testicle but not the lung, kidney and pancreas. Samples from the post mortem of a dog, which had to be destroyed for other reasons in a non-endemic area, were negative (Fig. 1). As described elsewhere, excess blood from tissues was removed by pre-blotting as blood contamination is known to interfere with probe sensitivity (McNerney et al. 1994).
Fig 1: post mortem examination of tissue samples from a Leishmania chagasi infected dog with radiolabelled Lmet2 DNA probe.
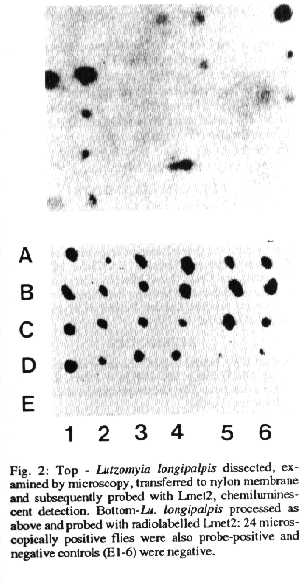
Lu. longipalpis fed on infected dogs - Xenodiagnoses were performed on dogs that had acquired natural infections of L. chagasi in Teresina and flies were examined 5-7 days after taking a blood meal. If flies were fed on skin with changes characteristic of canine VL they easily became infected (Table). Flies fed on apparently normal skin also became infected, although less frequently, confimfing the disseminated nature of the cutaneous infection in canine VL (Table). At day 5 after feeding, when gut contents and residual blood meals from dissected flies were transferred to membranes and probed, around 80% of microscopically positive flies were also probe-positive (Fig. 2, chemiluminescent probe). We have subsequently shown that protease treatment of blood-contaminated samples can restore sensitivity (McNerney et al. 1994) but chemiluminescent signals are still not necessarily obtained with such samples for flies in which very low numbers of organisms are seen by microscopy. At day 7, when there was less residual blood in the flies, 24 flies that were positive by dissection and microscopy were also positive with the DNA probe (Fig. 2, radiolabelled probe). The sensitivity of the Lmet2 probe with and without protocols to improve sensitivity for contaminated blood samples is discussed elsewhere (McNerney et al. 1994).
Fig. 2: Top - Lutzomyia longipalpis dissected, examined by microscopy, transferred to nylon membrane and subsequently probed with Lmet2, chemiluminescent detection. Bottom - Lu. longipalpis processed as above and probed with radiolabelled Lmet2: 24 microscopically positive flies were also probe-positive and negative controls (E1-6) were negative.
TABLE
Promastigote infection rates in Lutzomyia longipalpis 5-7 days after capture in kennels or after feeding on five naturally infected dogs
=============================================================
Transmissibility - Four sandflies, fed seven days previously on a naturally infected dog, were re-fed on a young dog from a non-endemic area: one of the four flies was shown subsequently by dissection and microscopy to have been infected at the time of the second feed. Within 22 days a lesion was present at the site of the bite (left inner ear). At 98 days after the infective feed the lesion had grown considerably, but apparently remained restricted to the inner ear. Interestingly, serology (indirect immunofluorescence and dot-enzyme-linked immunosorbent asssay) remained negative 98 days after infection and no amastigotes were found in bone marrow aspirates or spleen and liver biopsies. Other work has suggested that there may be a long prepatent period for seropositivity in canine VL (Ashford &: Bettini 1987, Dye et al. 1993). To check whether the cutaneous infection had disseminated away from the original site of infection, xenodiagnoses with colony-bred sandflies were performed on three separate occasions and on each occasion flies were fed at three sites - the left inner ear (on the lesion), the right inner ear and the abdomen. For flies fed on the left ear 48 out of 73 flies (66%) became infected (12/17, 23/39 and 13/17 on each occasion respectively). None of 108 and 79 flies fed respectively on the right ear and abdomen became infected, indicating that the infection had not yet disseminated from the original lesion. Clearly no general conclusions can be drawn from this one animal as to whether such ease of transmissibility dog-sandfly-dog is commonplace in Teresina but this would accord with the high prevalence of canine VL in the city: further experimental studies would clarify the course of infection in dogs and allow selection of the most appropriate means of parasitological and serological diagnosis.
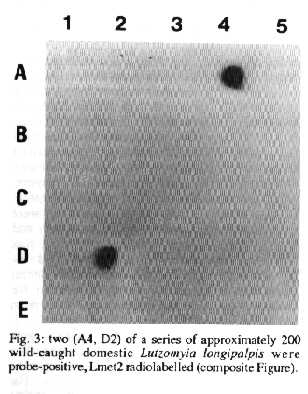
L. chagasi in wild-caught Lu. longipalpis - Dissection and microscopy of fed flies captured in a kennel with a sick dog subsequently shown to be infected demonstrated that a high proportion of such flies had promastigote infections (67%, Table). Infected flies were also found in a communal kennel with approximately 20 dogs which may or may not have been infected (Table). The Lmet2 DNA probe facilitates the screening of large numbers of sandflies for L. donovani-complex infectious and other studies have shown that sensitivity compares favourably with microscopy (Gebre-Michael el al. 1993). We have as yet examined a relatively small number (~200) wild-caught peridomestic and domestic Lu. longipalpis from Teresina by this method. Two infections were detected, both from sandflies captured in houses, confirming the potential usefulness of this procedure (Fig. 3).
Fig 3: two (A4, D2) of a series of approximately 200 wild-caught domestic Lutzomyia longipalpis were probe-positive, Lmet2 radiolabelled (composite Figure).
Overall these preliminary observations confirm the ease with which Lu. longipalpis can acquire infection from dogs with canine VL and suggest that the infection is highly transmissible dog-sandfly-dog. Similarly Rioux et al. (1979) have shown that L. infantum can be transmitted to a dog by a single sandfly. It is less certain that dog-sandfly-human is the main source of human VL (Evans el. al. 1992, Tavares et al. 1993) but experimental and circumstantial evidence suggest that this trausmission route is much more important than human-sandfly-human trausmission. In the Mediterranean form of human VL due to L. infantum and in human VL in Brazil blood parasitaemias are reported to be low (Ashford &: Bettini 1987, Lainson &: Shaw 1987) and Deane (1956) could only demonstrate blood parasitaemias in four of 43 symptomatic patients. Although sandflies can be infected by feeding on VL patients in Brazil and the Mediterranean region, Deane (1956) found that only four of 14 cases of human VL were infective to sandflies, whereas 12 out of 16 dogs were infective. Like humans, dogs have low numbers of circulating L. infantum or L. chagasi, but their efficiency as a source of infection is thought to be due to the abundant amastigotes in the skin. Amastigotes can be found in biopsies of apparently normal skin from patients with VL, but this is much less commonplace than in dogs. Thus Deane (1956) reported that five of 27 patients were positive by skin biopsy, and only one had abundant amastigotes, whereas 38 of 49 dogs were biopsy positive. Nevertheless in a recent study in State of Ceara, Vasconcelos et al. (1993) were able to recover L. chagasi from seven of 18 patients with proven kala-azar by culture of skin biopsies on Difco blood-agar. In contrast to L. chagasi/infantum, in Indian kala-azar due to L. donovani, large numbers of amastigotes can be found in the peripheral blood, many clinically ill patients can be shown to be parasitaemic and the infection is highly transmissable from humans to sandflies.
The Lmet2 DNA probe has proved to be a useful tool, especially for detecting infectious in sandflies. The value of the probe in its present form for the routine diagnosis of canine VL is uncertain: it seems unlikely to replace Giemsa-staining of skin biopsy impression or of bone marrow, spleen or popliteal lymph node aspirates except, perhaps, with further simplified procedures, if large numbers of animals are to be examined as part of widespread control programmes. Several rapid serological procedures are available, which are likely to be more practical for field use, although further comparative experimental studies are needed to determine how frequently they become positive early in the infection.
ACKNOWLEDGEMENTS
To Centre of Zoonoses in Teresina, particularly Vania Carvalho and Augusto Cesar, for their collaboraLion and support. To Dr Fernando Lima and Dr Amparo Salmito for general support from the Hospital for Infectious Diseases, Teresina. To Carlos HN Costa (Teresina), and Cesar Cuba Cuba (Brasilia) for helping to initiate collaboration between the Ministry of Health in Teresina and the London School of Hygiene and Tropical Medicine.
REFERENCES
Ashford RW, Bettini S 1987. Ecology and epidemiology: Old World, p. 365-424. In W Peters, R Killick-Kendrick (eds) The Leishmaniases in Biology and Medicine, Vol. 1.
Costa CHN, Pereira HF, Araujo MV 1990. Epidemia de leishmaniose visceral no Estado do Piaui, Brasil, 1980-1986. Rev Saude Publ 24: 361-372.
Deane LDM, 1956. Leishmaniose visceral no Brasil. Estudos sbre reservatorios e transmissores realizados no Estado do Ceara. Servico Nacional de Educacao Sanitaria, Rio de Janeiro, Brasil. 162pp.
Dye C, Vidor E, Demure J 1993 Serological diagnosis of leishmaniasis: on detecting infection as well as disease. Epidemiol Infect 103: 647-656.
Evans TG, Teixeira MJ, McAuliffe IT, Vasconcelos I, Vasconcelos AW, Sousa A de A Lima JW, Pearson RD, 1992. Epidemiology of visceral leishmaniasis in northeast Brazil. J Infect Dis 166: 1124-32.
Gebre-Michael T, Lane RP, Frame IA, Miles MA 1993. Leishmania donovani infections in phlebotomine sandflies from the kala-azar focus at Aba Roba in Ethiopia: DNA probe compared with conventional detection methods. Med Vet Entomol 7: 294-296
Howard MK, Kelly JM, Lane RP, Miles MA 1991. A sensitive repetitive DNA probe that is specific to the Leishmania donovani complex and its use as an epidemiological and diagnostic reagent. Mol Biochem parasitol 44: 63-72.
Howard MK, Ogunkolade W, Bryceson ADM, Davidson R, Moody All, Miles MA 1992. A DNA probe for human visceral leishmaniasis. Trans R Soc Trop Med Hyg 86: 35-36.
Lainson R, Shaw JJ 1987. Evolution, classification and geographical distribution, p. 1-120. In W Peters, R Killick-Kendrick (eds) The Leishmaniases in Biology and Medicine Vol. 1.
McNerney R, Frame IA, Vexenat J, Fonseca de Castro JA, Costa CHN, Siddiqui R, Dillon R, Wilson S, Miles MA 1994. Visceral leishmaniasis in Teresina, NE Brazil: towards a DNA probe kit and its adaptation to processing blood-contaminated samples. Arch Parasitol (in press).
Miles MA, Lainson R, Shaw JJ, Povoa M, De Souza AA 1981. Leishmaniasis in Brazil: XV. Biochemical distinction of Leishmania mexicana amazonensis, L braziliensis braziliensis and L braziliensis guyanensis - aetiological agents of cutaneous leishmaniasis in the Amazon Basin of Brazil. Trans R SocTrop Med Hyg 75: 524-529.
Rioux J-A, Killick-Kendrick R, Leaney AJ, Young CJ, Turner DP, Lanotte G, Bailly M 1979. Ecologie des leishmanioses dans la sud de la France 11. La leishmaniose viscrale canine - success de la transmission experimentale "Chien-Phlebotome-Chien" par la pirure de Phlebotomus ariasi. Ann Parasitol Huma Comp 54: 401-407.
Tavares, JP, Alves Pereira FC, Fonseca de Castro JA, Goldin A, Siddiqui R, Vexenat A, Furtado Campos JD, Pereira HF, Lima FC, Costa CHN, Miles MA 1993. The distribution of visceral leishmaniasis in the suburban focus of Teresina, State of Piaui, Brazil. Mem Inst Oswaldo Cruz 88 Supl. 235.
Vasconcelos AB de, Sousa A de Q, Vasconcelos AW, Diogenes MJN, Momen H, Grimaldi G, Menezes DB, Sleigh AC 1993. Parisitisme cutan par Leishmania (Leishmania) chagasi au cours de la leishmaniose viscrale Sud-Americaine. Bull Soc Path Ex 86: 101-105.
Wilson SM, McNerney R, Moreno MB, Frame I, Miles MA 1992. Adaptation of a radioactive L. donovani complex DNA probe to a chemiluminescent detection system gives enhanced sensitivity for diagnostic and epidemiological applications. Parasitology 104: 421-426. Copyright 1994 Fundacao Oswaldo Cruz - FIOCRUZ The following images related to this document are available:Halftone images[oc94028b.gif] [oc94028a.gif] [oc94028c.gif]Photo images[oc94028b.jpg] [oc94028c.jpg] [oc94028a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}