 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Comparative Evaluation of a Simple and Sensitive Assay for Detection of Orthomyxo and ParamyxovirusesJNSS Couceiro (+), ESS Couceiro (*), AMV Pinto (**), RD Machado, MC Cabral
Code Number: OC94038
Size of Files:
Text: 21K
Graphics: Line Drawing (Gif) - 11K
Departamento de Virologia, Instituto de Microbiologia, CCS, Bloco I, Cidade Universitaria, Caixa Postal 68040, 21941-590 Rio de Janeiro, RJ, Brasil
Studies were done to evaluate comparatively the traditional HA assay and a more recently introduced lectin-neuraminidase (LN) methodology in search of a simple and sensitive assay for virus detection during laboratorial diagnosis. The results proved the value of LN assay as a sensitive methodology for detection of virus particles, presenting results at least equal to those obtained by HA (hemagglutination) assay, with significant values of accumulated frequencies for LN/HA factors (ratios between LN and HA titers) higher than two. The accumulated values of frequencies for LN/HA factors as high as four were very significant, 72.7% for influenzavirus and 60.7% for Newcastle disease virus (NDV), moreover accumulated frequencies for LN/HA factors even as high as 32 were due to influenzavirus (45.4%) and NDV (7.2%) samples. After the storage period, most of those concentraded samples that even did not present HA titers could be detected through LN assay, demonstrating a lower threshold for virus detection.
Key words: hemagglutination assay - lectin-neuraminidase assay - influenzaviruses - parainfluenzaviruses
Influenzaviruses and Newcastle disease viruses (NDV), classified as orthomyxovirus and paramyxovirus respectively, are RNA-viruses that presents capsid with helical symmetry and lipoprotein envelope, where glycoprotein structures exhibit adsorption activity for cell receptors besides sialidase and fusion activities. In influenzaviruses, the hemagglutinin (HA) trimeric structure is involved in adsorption and fusion processes, which are expressed by hemagglutination and hemolysis when using erythrocytes as target cells, while the neuraminidase (NA) structure is responsible for sialidase cleaving activity on cell receptors (Murphy & Webster 1990). In NDV, the HN structure is responsible for adsorption (hemagglutinating) and sialidase activities, while the F structure is involved in the fusion (hemolysis) process (Kingsbury 1990).
N and HN spikes that present sialidase activity, when inserted in cellular cytoplasmatic membranes or virus envelopes, develop hydrolysis of sialic acids residues on carbohydrated chains of membrane receptors, which change to exhibit galactose molecules as terminal residues (Baum & Paulson 1991).
Since Hirst (1941) and McClelland and Hare (1941) classical descriptions about the fast hemagglutination process, most of the researches done on orthomyxoviruses and paramyxoviruses have used their hemagglutinating function as the analysis parameter to quantify the virus preparations. The standard method for the study of virus sialidase activity have not been widely used, which could be explained by the low paractibility of methodologies such as Warren test (Warren 1959), which reveals enzimatically the sialic acid residues released through the virus sialidase activity. This lectin- neuraminidase (LN) assay was as easily developed as the conventional hemagglutination assay, the former exhibits advantage over that traditional HA methodology, showing practibility and great sensitivity. In this technique, after the cleavage activity on sialic acids residues expressed by the virus sialidase, those galactose residues exposed on the carbohydrate chains of cell receptors are revealed by addition of peanuts agglutinin (PNA). The reaction between the PNA tetrameric structure and galactosyl end-groups exposed on erythrocytes membranes originates a hemagglutination reaction, revealing finally the presence of virus particles through their sialidase activity (Lierner et al. 1986).
This study, utilizing a significant number of samples, was done to compare the efficiency of the HA and LN assays, as techniques for virus titration and diagnosis. The experiments intended also to establish the ideal conditions for reaction.
MATERIALS AND METHODS
Virus samples - Eleven clinical and standard samples of human and avian influenza A (H1N1, H6N2 and H7N1) viruses and 28 clinicala and standard samples of NDV were analyzed as shown in Tables I and II, totalizing 39 samples. NDV standard samples of low (lentogenic) and high (velogenic) pathogenic potential were used.
Preparation, concentration and storage for different periods of time - Standard virus samples for this study were prepared by inoculation into allantoic cavity of 10-day old embryonated chicken eggs and incubation at 36 oC for 48 hr (Rovozzo & Burke 1973). Clinical samples were considered as those detected by hemagglutination assay after one or two passages of clinical materials from birds (feces) into allantoic cavity, under similar incubation conditions. After incubation, harvested allantoic fluids were clarified by centrifugation at 7,500x g for 30 min at 4 oC and tested by hemagglutination assay (Sever 1962). The clinical samples were also typed by hemagglutination inhibition assay (Sever 1962). The hemagglutinating virus samples were finally 50x concentrated by centrifugation at 100,000x g for 60 min. The pellets from the virus samples were resuspended, with TESC pH 8.4 (0.01 mM TRIS, 0.01 mM EDTA, 0.10 M sodium cloride, 0.006 mM cystein) and stored for different periods of time at -20 oC until utilization (Rovozzo & Burke 1973). Non concentrated standar virus samples of influenzaviruses A H3N2 and parainfluenzaviruses type 1 (Sendai) were used for additional studies.
Erythrocytes - Human O group erythrocytes were used after collecion in Alsever solution, three washing steps and suspension at 10% concentration with 0.15 M sodium chloride. The final suspension at 1% concentration was prepared with 0.15 M phosphate buffer saline (PBS) pH 7.2. In preliminary experiments due to establish the ideal pH for reaction and optimal period of time for incubation, different buffers (Maeda & Ohnishi 1980) were used to prepare 1% erythrocytes at different pH values.
Lectin - PNA (peanuts lectin/Arachis hypogaea) obtained from Sigma, was used after titration and standardization at 25 oC to two hemagglutinating units (Lothan et al. 1975).
Hemagglutination Assay (HA) - Triplicates of virus samples (25 ul) were serially diluted in 25 ul volumes of 0.15 M PBS pH 7.2, with posterior addition of 25 ul volumes of 1% human O group erythrocytes prepared in 0.15 M PBS pH 7.2 or in buffers at different pH values, 0.2% bovine albumin was also added. The results were read after incubation for 2 hr at 4 oC. The reciprocal of the highest dilution (geometric mean of the triplicates) of each virus sample causing complete agglutination was considered as its HA titer per 25 ul (Sever 1962).
Lectin-Neuraminidase assay (LN) - Triplicates of virus samples (25 ul) were serially diluted with 25 ul volumes of 0.15 M PBS pH 7.2 and 1% human O group erythrocyte suspensions prepared in 0.15 M PBS pH 7.2 or in buffers at different pH values were added, 0.2% bovine albumin was also added. The reactions were incubated at 4 oC (negative controls) or at 37 oC for 2 hr until complete reversion of the initial hemagglutination due to virus binding, when PNA was added after homogenization of the wells that presented complete reversion of the initial hemagglutination. Cell controls with (erythrocytes + buffer + lectin) and without lectin (erythrocytes + buffer) wer also prepared. The LN titer (per 25 ul) of each virus sample was considered as the reciprocal of the highest virus dilution (geometric average) which was causing complete agglutination for PNA, after 1 hr of incubation at 25 oC (Luther et al. 1983).
HA and LN assays. Preliminary analysis to establish ideal pH value for reaction and optimal period of time for incubation - Influenza A H3N2 Virus (A/England/42/72) sample was analyzed by HA (4^oC) and LN (37^oC) assays, at pH values ranged from 5.0 to 9.0, using acetate, sodium phosphate and glycine-NaOH buffers as recommended by Maeda and Ohnishi (1980). Experiments were developed in search of the optimal pH value to be used in HA and LN assays and the period of incubation necessary for complete reversion of the initial hemagglutination, before addition of PNA.
HA and LN assays. Extensive analysis using pH and incubation parameters established as above - Clinical and standard concentrated samples of influenzaviruses and NDV (Tables I, II) were studied by HA and LN assays at pH value and for period of time such as established above. Unconcentrated standard influenzaviruses (A/PR/8/34 H1N1, A/England/42/72 H3N2) and parainfluenzavirus type 1 (Sendai) samples were studied in an additional analysis. The LN/HA Factor that expresses the advantage of the LN assay over the conventional HA assay was calculated as:
virus titer in LN assay
LN/HA factor = -----------------------
virus titer in HA assay
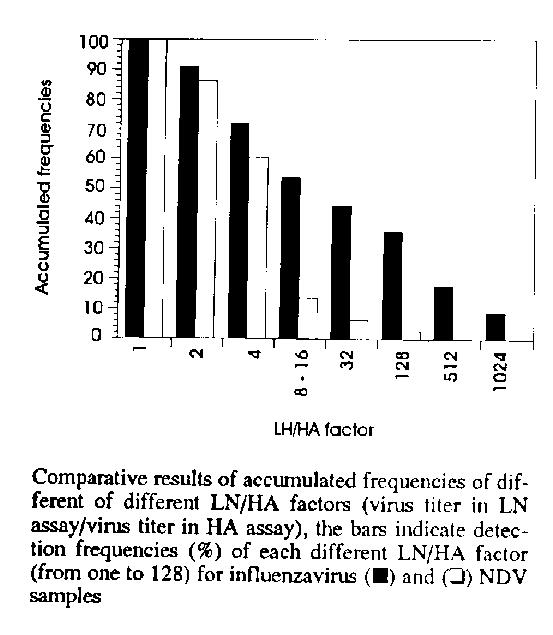
The data from this analysis on samples of influenzaviruses and NDV were also expressed (Fig.) through decreasing values of accumulated frequencies due to gradually increasing LN/HA factors.
RESULTS AND DISCUSSION
TABLE I
Comparative analysis between titers obtained through hemagglutination (HA) and lectin-neuraminidase (LN) assays for clinical and standard influenzavirus samples
======================================= Influenza HA LN LN/HA sample (a) titer titer factor --------------------------------------- Clinical 2 2 1 A/H7N1 Standard A/PR/8/34 16 32 2 (H1N1) Clinical 4 8 2 A/H7N1 Clinical 32 128 4 A/H7N1 Clinical 2 8 4 A/H6N2 Clinical 32 512 16 A/H7N1 Sumdard A/PR/8/43 8 256 32 (H1N1) Clinical < 2 128 128 A/H7N1 Clinical < 2 128 128 A/H7N1 Clinical < 2 512 512 A/H7N1 Clinical < 2 1024 1024 A/H7N1 ====================================a: clinical samples typed by hemagglutination inhibition assay (Sever 1962). Standard influenzavirus samples were kindly sent by Dr JJ Skehel (National Institute for Medical Research, Mill Hill, London).
TABLE II
Comparative analysis between titers obtained by hemagglutination (HA) and lectin-neuraminidase (LN) assays for clinical and slandard NDV samples
================================================================ NDV (a) HA LN LN/HA NDV HA LN LN/HA sample titer titer factor sample titer titer factor ---------------------------------------------------------------- Standard Standard lentogenic 2 2 1 lentogenic 4 16 4 B1 Lasota Standard Standard lentogenic 64 64 1 velogenic 64 256 4 B1 SO-93 Standard lentogenic 4 4 1 Clinical 2 8 4 B1 Standard lentogenic < 2 < 2 1 Clinical 2 8 4 B1 Standard lentogenic 16 32 2 Clinical 2 8 4 Standard lentogenic 8 16 2 Clinical < 2 8 8 Standard lentogenic 4 8 2 Clinical < 2 8 8 Standard lentogenic 16 32 2 Clinical 4 32 8 Standard lentogenic 4 8 2 Clinical 8 64 8 Standard lentogenic 8 16 2 Clinical < 2 16 16 Standard lentogenic 2 4 2 Clinical 16 256 16 Standard lentogenic 4 16 4 Clinical < 2 32 32 Standard lentogenic 2 8 4 Clinical 8 256 32 Standard lentogenic 2 8 4 Clinical < 2 128 128 =============================================================== a: clinical samples typed by hemagglutination inhibition assay (Sever 1962). Lentogenic (B1 and La Sota) and velogenic (SO-93) standard NDV samples were kindly sent by Dr AM Souza (Empresa Brasileira de Agropecuaria, Rio de Janeiro Brazil).
The preliminary analysis determined the pH 7.2 as an intermediate value of neutral pH at which suspensions of erythrocytes should be adjusted for LN assay, as traditionally used for HA methodology (data not shown). Low pH values (5.0-5.2) were responsible for hemolysis while pH values higher than 7.8 did not produce significant LN/HA factors (> 2). This analysis was done with an influenzavirus A/England/42/72 sample, where a period of 3 hr and 30 min at pH 7.2 was also established as necessary for a complete reversion of an initial HA reaction.
The analysis procedures were developed at pH 7.2 as determined above, using the same diluent (PBS) recommended by World Health Organization for biochemical evaluation of the sialidase activity (Luther et al. 1980, 1983). The results obtained by comparative analysis of 39 lentogenic and velogenic NDV and influenzaviruses (H1N1, H6N2, H7N1) samples through agglutinating titers obtained in HA and LN assays, can be observed in Tables I and II. The LN/HA factors (relation between LN and HA titers) were expressed in those tables in relation to influenzavirus and NDV samples. The accumulated values of frequencies for LN/HA factors equal to four were very significant, 72.7% (influenzavirus) and 60.7% (NDV), moreover accumulated frequencies of factors as high as 32 for influenzavirus (45.4%) and NDV (7.2%) samples were found (Fig.). In relation to influenzavirus samples, significant values of accumulated frequencies even for LN/HA factors as high as 512 (18.1%) and 1,024 (9.0%) were observed.
The results proved the value of LN assay as sensitive and simple methodology for detection of virus particles, presenting results at least equal to those observed in HA assay, with high values of accumulated frequencies for LN/HA factors higher than two. The efficiency of the technique as a new methodology to be used for virus diagnosis was evident. After different storage periods at 20 oC, even those concentrated standard and clinical samples that did not present titers by HA assays could be detected by the LN assay, demonstrating a lower threshold for virus detection. The same efficiency was also demonstrated when using only unconcentrated virus samples (data not shown).
In this study, the analysis indicated the importance of this simple and sensitive methodology for detection of virus particles, in diagnosis of diseases, showing advantage even over the traditional HA assay. This LN assay have exhibited good sensitivity when compared to Warren test and showed to be a practical methodology to reveal the virus sialidase activity, as already indicated above (Luther et al. 1983, Fiszon & Hannoun 1990). McNulty and his group (McNulty et al. 1975) tried to associate the sialidase content and the virulence of velo, meso and lentogenic samples of NDV, aspect that has been explored by other researches (Schulman & Palese 1977, Ogawa & Ueda 1981).
This technique may play an important role for studies on the sialidase activity, which is also present in many other microorganisms, such as trypanosomatides (trans-sialidase) and diverse genus, with comparative results already demonstrated by Warren test, a traditional enzymatic methodology (Pereira 1983, Breighton & Whiley 1990, Heuermann et al. 1991, Briselden et al. 1992). This method has also been used in the studies on processes of cellular interactions (Peacock et al. 1990).
ACKNOWLEDGEMENTS
To Mr Joao Rugel for technical assistance and Mrs Elizabeth Brito Ferrarezzi for office facilities.
REFERENCES
Baum LG, Paulson JC 1991. The N2 neuraminidase of human influenza virus has acquired a substrate specificity complementary to the hemagglutinin receptor specificity. Virology 180: 1015.
Breighton D, Whiley RA 1990. Sialidase activity of the "Streptococcus milleri group" and other viridans group streptococci. J Clin Microbiol 28: 1431-1433.
Briselden AM, Moncla BJ, Stevens CE, Hillier SL 1992. Sialidases (neuraminidases) in bacterial vaginosis and bacterial vaginosis- associated microflora. J Clin Microbiol 30: 663-666.
Fizson B, Hannoun C 1990. Comparison of neuraminidase of the same subtype but from different species using a new method of titration. J Virol Methods 27: 79-90.
Heurmann D, Roggentin P, Kleineidam RG, Schauer R 1991. Purification and characterization of a sialidase from Clostridium chauvoei. Glyconjugate J 8: 95-101.
Hirst GK 1941. Agglutination of red cells by allantoic fluid of chick embryos infected with influenza with influenza virus. Science 94: 22-23.
Kingsbury DW 1990. Paramyxoviridae and their replication. p. 945- 962. In BN Fields, DM Knipe (eds), Virology, 2nd ed., Raven Press, New York.
Lierner IE, Sharon N, Goldstein IJ 1986. The Lectins. Properties, functions and applications in Biology and Medicine. Academic Press Inc., Harcourt Brace Jovanovich Publishers, New York.
Lothan R, Skutelsky E, Danon DJ, Sharon N 1975. The purification, composition and specificity of the anti-T-lectin from peanut (Arachis hypogaea). J Biol Chem 150: 8518-8523.
Luther P, Adamczyk B, Bergmann K 1980. Simple test for detection of virus neuraminidase and anti-neuraminidase using lectins (Lectin-Neuraminidase Test System). Zbl Bakt Hyg I, Abt Orig A 248: 281-285.
Luther P, Klett GE, Weber S, Pechmann H, Bergmann KCH 1983. The lectin Neuraminidase Inhibition Test: A new method for the detection of antibodies to neuraminidase. J Biol Stand 11: 115-121.
Maeda T, Ohnishi S 1980. Activation of Influenza Viruses by acidic media causes haemolysis and fusion of erythrocytes. FEBS Letters 122: 283-287.
McClelland L, Hare R 1941. The adsorption of influenza virus by red cells and a new in vitro method of measuring antibodies for influenza virus. Canad J Publ Hlth 32: 530-538.
McNulty NS, Gowans EJ, Houston MJ, Fraser G 1975. Neuraminidase content of strains of Newcastle disease virus which differs in virulence. J Gen Virol 27: 399-402.
Murphy BR, Webster RG 1990. Orthomyxoviruses, p. 1091-1152. In BN Fields, DM Knipe (eds) Virology, 2nd ed. Raven Press, New York.
Ogawa T, Ueda M 1981. Genes envolved in the virulence of an avian influenza virus. Virology 113: 304-313.
Peacock JS, Colsky AS, Pinto VB 1990. Lectins and antibodies as tools for studying cellular interactions. J Immun Methods 126: 147-157.
Pereira MEA 1983. A developmentally neuraminidase activity in Trypanosoma cruzi. Science 219: 1444-1446.
Rovozzo GC, Burke CN 1973. A Manual of Virological Basic Techniques. Prentice-Hall Inc., Engle-wood Cliffs, New Jersey.
Schulman J, Palese P 1977. Virulence factors of influenza A viruses: WSN virus neuraminidase required for plaque production in MDBK cells. J Virol 24: 170-176.
Sever JL 1962. Application of a microtechnique to viral and serological investigations. J Immunol 88: 320-329.
Warren L 1959. The thiobarbituric acid assay of sialic acids. J Biol Chem 234: 1971-1975.
Copyright 1994 Fundacao Oswaldo Cruz - FIOCRUZ The following images related to this document are available:Line drawing images[oc94038a.gif] |
| |||||||||

{kind=link}