 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
An Inhibition ELISA to Determine Alpha Macroglobulin Levels in Mouse PlasmaMauricio RMP Luz (+), Fred Van Leuven (*) , Tania C Araujo-Jorge
Code Number: OC94041
Size of Files:
Text: 19K
Graphics: Line Drawings (Gif) - 44.3K
Departamento de Ultraestrutura e Biologia Celular, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil * Dept. of Human Genetics, Katholic University of Leuven, Gasthuisberg, Herestraat 49, Leuven B-3000, Belgium
A sensitive method for quantifying mouse plasma alpha- macroglobulins (AM) using an inhibition ELISA is described. AM are important plasma proteinase inhibitors that possibly act also as immunomodulatory molecules. The standard protocol developed in our experiments involves coating well with 10ug/ml A2M in carbonate buffer, followed by incubation with a 1:1 (v/v) mixture of the plasma to be tested (diluted 1/1000) and goat anti-AM (diluted 1/1250). This is followed by further incubation, first with the enzyme-conjugated antibody and with the substrate prior to the reading of absorbance levels of the reaction products. Standard curve samples must be included in each plate, employing known amounts of the purified Murine Alpha-2-Macroglobulin (MuA2M) used for coating, with concentrations ranging from 0.001 to 10 ug/ml. Using test samples in triplicates and a 6-point standard curve in a single ELISA plate, 25 plasma samples can be tested accurately. The method offers an useful tool for establishing AM levels in small samples of mouse plasma.
Key words: alpha-2-macroglobulin - A2M - ELISA - murinoglobulin
Plasma proteinase inhibitors are the necessary counterparts of proteinases that participate in many physiological and pathological processes. Alpha-macroglobulins (AM) form a family of especially important plasma proteinase inhibitors, since they are able to inactivate eucariotic and procariotic endopeptidades of all classes (serine, cysteine, aspartic acid and metallo-type proteinases), acting like "molecular traps" for proteinases (Sottrup-Jensen 1989). In addition to their role in the clearance of proteinases, AM have been proposed as a second line regulatory element in extravascular proteolysis (Van Leuven 1982), and have also been implicated in immunomodulatory events (La Marre et al. 1991).
All mammals contain one or more AM some of which are expressed constitutively, while others are inducible in inflammation (La Marre et al. 1991). Rodents have multiple members of the AM family. In mice, two forms of AM are known, the constitutive tetrameric alpha-2-macroglobulin, homologous to human A2M (MuA2M), and a monomeric form called murinoglobulin (MuG), homologous to rat alpha 1-protein inhibitor 3. MuG exhibits the molecular mechanism for proteinase trapping typical of the AM family, and shows antigenic similarities with MuA2M (Abe et al. 1989). A detailed study of the MuG has revealed the existence of genes coding for proteins that are not found in plasma, at least in normal situations (Overbergh et al. 1991).
Methods to detect and quantify AM have been described mainly for human proteins (James et al. 1966, Ganrot & Schersten 1967, Polvesen et al. 1987, Petersen et al. 1985). However, immunodiffusion (Tunstall et al. 1975) and rocket immunoelectrophoresis (Issac et al. 1990) have also been used to quantify mouse A2M, using an anti-MuA2M serum prepared in the laboratory. In quantitative enzyme-linked immunoassays (ELISA) it is usually necessary to combine two antisera against the antigen to design a sandwich ELISA (Petersen et al. 1985, Polvsen et al. 1987, Banks et al. 1990). In a variant on this direct method, antigens can be quantified in an inhibition ELISA using only one antiserum (Carlier et al. 1978). In this paper we describe a sensitive method for quantifying mouse plasma AM using an inhibition ELISA.
MATERIALS AND METHODS
Mice - Outbred Swiss albino mice were obtained from the animal facilities of FIOCRUZ (Rio de Janeiro, Brasil). Eight week old animals were used throughout the experiments.
Blood collection and processing - 30-40 ul of blood were collected on heparin, either by cardiac puncture or from the tail using micro-hematocrit capillaries. Plasma samples were kept at -20 oC until analysis.
Isolation of murine A2M - MuA2M was isolated from a pool of freshly obtained mouse plasma as previously described (Overbergh et al. 1991). Briefly, the plasma were treated with BaCl and BaSO, and MuA2M precipitated between 4% and 8% (w/v) of polethylene glycol 6000. The precipitate was solubilized and dialyzed against PBS and fractionated on a column (5 x 90 cm) of Ultrogel ACA22 (LKB, Sweden). Eluent was a sodium phosphate buffer (25 mM, pH 7.7). The final purification step consisted of affinity chromatography on a column (2.6 x 90 cm) of Blue Sepharose (Pharmacia, Sweden). The purity of the different AM fractions eluted was analyzed by rate electrophoresis and by SDS-PAGE. Purified MuA2M was used to coat ELISA plates and to make standard curves.
Anti-MuA2M immune sera - Goat anti-MuA2M serum was obtained from Nordic Immunologicals (Holland), and its specificity was tested by immunoprecipitation (IP) in agarose gels or by Western blotting with total mouse plasma and purified MuA2M and MuG (Van Leuven et al. 1987).
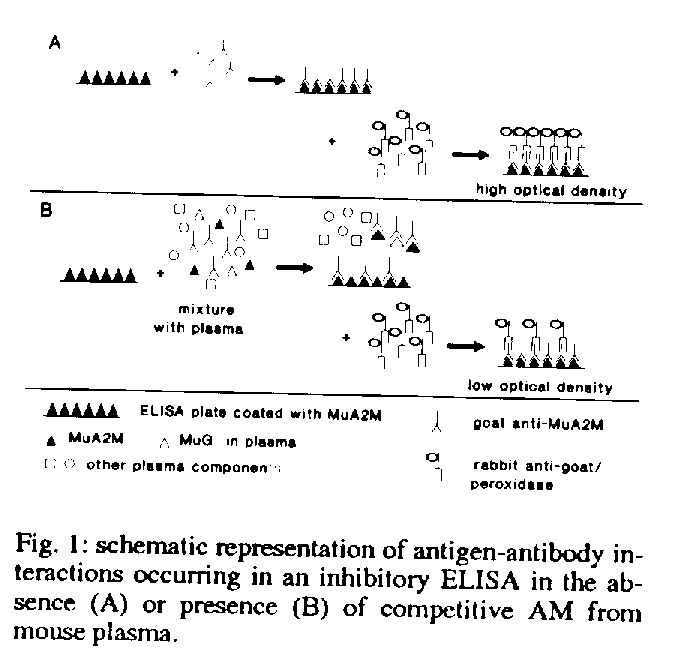
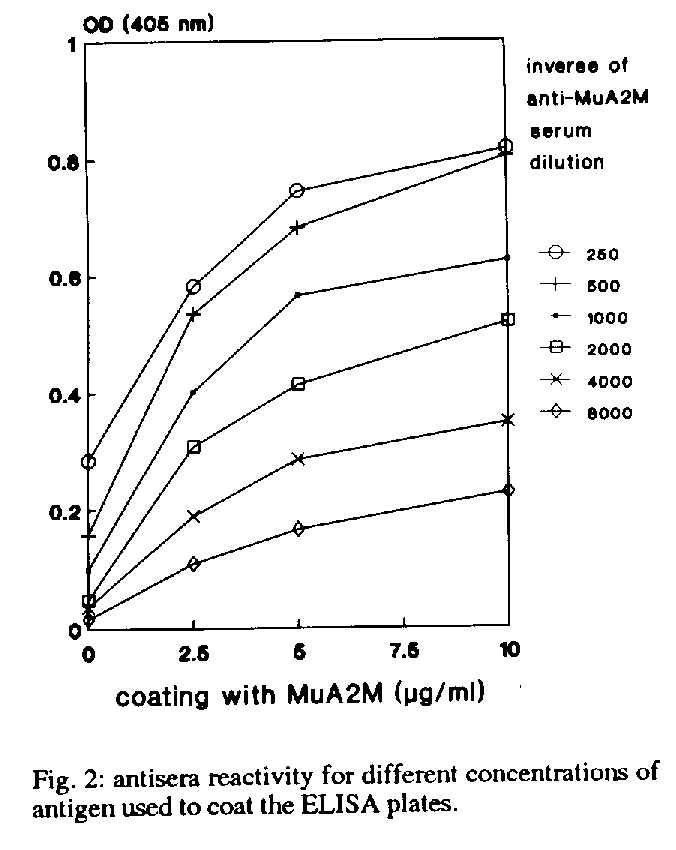
Alpha-macroglobulin quantification - 96-well immunoplates were coated overnight with 50 ml (0.5 mg) of purified MuA2M in 0.1 M NaHCO3 buffer pH 9.6. The remaining reactive sites were blocked with 1% BSA (1 hr) prepared in the same buffer and afterwards the plates were washed three times in 0.01M phosphate buffer pH 7.2 containing 0.15 M NaCl and 0.5% Tween 20 (PBS-T). Anti-MuA2M goat sera were previously mixed in eppendorf tubes, either with the murine plasma to be tested or with standard solutions of MuA2M (ranging from 0.3 to 10 ug/ml), and incubated for 1 hr at room temperature in a rotatory shaker. This mixture was added to the MuA2M coated wells and incubated for 2-4hr at 37 oC. After 3 PBS-T washes the plates were incubated with rabbit anti-goat IgG peroxidase conjugated (Sigma Chemical Co.). Enzymatic activity was measured at 450 nm using H2O2 and 3,3',5,5'- tetramethylbenzidine (TMB) as chromogenic substrate. The AM concentration in each sample was calculated from the standard curve prepared by linear regression from the logarithm of the optical densities obtained in each plate (see results for details). Results are expressed in mg/ml. Fig. 1 shows a diagram of the inhibition ELISA assay. Figure 2 antisera reactivity for different concentrations of antigen used to coat the ELISA plates Figure 3 levels of reactivity of anti-MuA2M binding to ELISA plates coated with 10 ug/ml. Figure 4 a typical standard curve obtaines using plates coated with 10 ug/ml MuA2M,, anti-MuA2M 1/2000 and peroxidase conjugated 1/2000
RESULTS
Determination of optimal concentrations of antigens and antibodies - For the inhibition ELISA it is crucial to determine the optimal relationship - in terms of relative quantities - between the antigen bound to the plates (MuA2M) and the antibody in solution (anti-MuA2M). All the specific antibodies must bind to the antigen on the plate, in such way that small amounts of soluble MuA2M (added to compete with the antigen in the coat) are sufficient to reduce the quantity of antibody available to bind to the plate. This decrease on antibody availability leads to a proportional decrease in the final optical density obtained after incubation with appropriate system of revelation (Fig. 1). To avoid undetected consumption of antibodies, they should not be present in excess.
Experiments in which ELISA plates were coated with purified MuA2M solutions at different concentrations (from 2.5 to 10ug/ml), saturated with BSA and further incubated with anti-MuA2M also at different dilutions, are shown in Fig. 2. Coating the plates with 5-10 ug/ml allowed the detection of MuA2M using anti-MuA2M dilutions of up to 1/2000 (Fig. 2) andieldedoptical densities higher than 0.5. So a concentration of 10 mg/ml MuA2M was considered to be optimal for the coating of plates, and was used thereafter. The binding of anti-MuA2M to the antigen-coated plates increased in a dose dependent manner, and high levels of reactivity were obtained at the dilution of 1/500 (Fig. 3). Sub-saturation levels, such as 1/1000 or 1/2000, were found to be optimal. Standard curves constructed using such dilutions showed that at the higher dilution a better slope could be obtained (Fig. 3, inset). A typical standard curve is shown in Fig. 4. The optimal sub-saturation level of the anti-MuA2M had to be titrated for each batch of serum, in order to reproduce sensitive standard curves; appropriated dilutions were generally found to be in the range of 1/1000 to 1/3000 (data not shown).
TABLE I
Reproducibility of ELISA using different batches of the reagents or different dilutions
=============================================
Exp. MuA2M anti- peroxidase OD
10 ug/ml MuA2M conjugate (405nm)
(coating) 1/500
---------------------------------------------
1 batch 1 batch 1 1/2000 0.780
batch 2 0.772
batch 3 0.683
1 batch 2 batch 1 0.723
batch 2 0.738
batch 3 0.650
2 batch 1 batch 2 1/1000 1.579
1/2000 1.488
1/5000 0.715
=============================================
Obs: peroxidase substrate used in all cases: TMB
TABLE II
ELISA reproducibility with different plasma samples
===================================================
Sample Exp (a) AM (mg/ml) Mean (sd) (c)
---------------------------------------------------
I a 1.15 1.36 1.32(b) 1.28 (0.11)
b 1.13 1.05 1.37 1.22 (0.13)
II a 2.96 2.74 3.13 2.94 (0.20)
b 2.05 3.08 2.83 2.65 (0.44)
III a 4.00 3.64 3.90 3.85 (0.31)
b 3.44 4.20 4.16 3.93 (0.35)
IV a 3.53 2.30 3.09 3.31 (0.31)
b 3.63 3.79 4.46 3.93 (0.46)
===================================================
a: indicates two different expenments in which the levels of
AM of a same sample were determined b: triplicates in a same plate c: standard deviation
Reproducibility of the assays - Since this inhibition ELISA was designed to depend only on the availability of the purified protein and one antiserum, it was essential to determine the va-riability of the assay when using different batches of these reagents. Table I shows that using two batches of purified MuA2M and three batches of commercial goat anti-MuA2M we obtained similar optical density levels. It was important to titrate the peroxidase conjugate, since otherwise it might have given variable absorbance readings (Table I).
Reproducible results were also obtained using different plasma samples (Table II), thus suggesting that the assay is also efficient for the detection of AM in complex mixtures.
DISCUSSION
Proteinases are powerful enzymes that can activate or inactivate target proteins, as well as participate in their maturation or destruction, by cleaving some of their peptide bonds. Since AM are among the most diverse and abundant plasma proteinase inhibitors, their levels may indicate the load of proteinases at any given moment in an organism. A search through the literature revealed that, to date, all the methods used to quantify mouse AM have involved immuno-difusion (radial or on a electrophoretic field), despite the fact that enzyme linked immunosorbent assays, as well as being more sensitive than immuno-precipitation techniques, are also easier and faster to perform. In the present paper, we have des-cribed an inhibition ELISA that can be used to quantify the total amount of AM in mouse plasma. The method uses reagents that, for the most part, are commercially available, such as goat anti-mouse A2M, peroxidase-conjugated rabbit anti-goat IgG and peroxidase substrate. The only laboratory-prepared reagent is purified murine AM. Since A2M is easy to isolate and purify, by simple methods such as affinity chromatography or gel filtration (Virca et al. 1978, Kurecki et al. 1979, Van Leuven et al. 1985, Shibuya et al. 1988), this is not a limiting factor.
The standard protocol used in our experiments involves coating wells with 10 ug/ml A2M in carbonate buffer, blocking them with 1% BSA and incubating them with a 1:1 (v/v) mixture of the plasma to be tested (diluted 1/1000) and goat anti-AM (diluted 1/1250). This is followed by further incubation, first with enzyme-conjugated antibody and then with substrate, prior to the reading of the absorbance levels of the reaction product. Standard curve samples must be included in each plate, employing known amounts of the purified used for coating, with quantities ranging from 0.001 to 10 ug/ml. Thus using test samples in triplicate and a 6-point standard curve, in a single ELISA plate 25 plasma can be tested accurately.
It is important to point out that, in contrast to gel immunoprecipitation techniques, in which lines of precipitation are formed monospecifically by the cross-linking of antigen and antibodies, in ELISA all antigens epitopes can be recognized by antibodies in polyclonal antisera. This contributes to the enhanced sensitivity of the method. No monoclonal antibodies are available for murine AM, but, in view of the high structural homologies among different AM proteinase inhibitors, polyclonal antisera against heterologous A2M and MuG can be used to detect the total circulant AM, without discrimination between A2M and MuG. In studies using rocket immuno-lectrophoresis, A2M and MuG have each been found to occur in mouse plasma under normal conditions at concentrations between 0.5 and 1 mg/ml each one (Overbergh et al. 1991). We therefore expected the overall level of AM to be in the region of 1-2 mg/ml. This was the case in our ELISA tests on plasma from Swiss mice.
The method described in this paper may offer a useful tool for establish AM level in mice, and for tracking variations in AM concentrations under different physiological and pathological conditions. We have already used it successfully to show that enhanced AM level in BALB/c mice infected with Trypanosoma cruzi correlate with resistance to acute infection (Araujo-Jorge et al. 1992).
ACKNOWLEDGMENTS
To Yves Carlier and Carine Truyens for helpful discussions at the outset of this study.
REFERENCES
Abe K, Yamamoto K, Sinohara H 1989. Proteinase inhibitory spectrum of mouse murinoglobulin and alpha-macroglobulin. J Biochem 106: 564-568.
Araujo-Jorge TC, Lage MJF, Rivera MT, Carlier Y, Van Leuven F 1992. Trypanosoma cruzi: enhanced alpha-macroglobulin levels correlate with resistance of BALB/cj mice to acute infection. Parasitol Res 78: 215-221.
Banks RE, Evans SW, Van Leuven F, Alexander D, McMahon MJ, Whicher JT 1990. Measurement of the "fast" or complexed form of alpha-2-macroglobulin in biological fluids using a sandwich enzyme immunoassay. J Immunol Methods 126: 13-16.
Carlier Y, Bout D, Fruchart JC, Desreumax C, Dewailly P, Sezile A, Jaillard J 1978. Inhibition enzyme-immunoassay. Application to human apolipoprotein B. J Immunol Methods 21: 317-324.
Ganrot PO, Schersten B 1967 Serum alpha-2-macroglobulin concentration and its variation with age and sex. Clin Chim Acta 15: 113-120.
Isaac L, Pereira M, Santos M, Sampaio EP, Lima NR, Laje MJF, Araujo-Jorge TC 1990. Trypanosoma cruzi: plasma levels of alpha-2-macroglobulin during experimental murine infections with reticulotropic and myotropic strains. Parasitol Res 76: 726-728
James K, Johnson G, Fudenberg HH 1966. The quantitative estimation of alpha-2-macroglobulin in normal, pathological and cord sera. Clin Chim Acta 14: 207-214.
Kurecki T, Kress LF, Laskowski M 1979. Purification of human plasma alpha-2-macroglobulin and alpha-1 proteinase inhibitor using zinc chelate chromatography. Anal Biochem 99: 415-420.
La Marre J, Wollenberg GK, Gonias SL, Hayes MA 1991 Biology of disease: cytokine binding and clearance properties of proteinase-activated alpha-2-macroglobulin. Lab Invest 65: 3-8.
Overbergh L, Torrekens S, Van Leuven F, Van Der Berghe H 1991. Molecular characterization of murinoglobulins. J Biol Chem 254: 16903-16910.
Petersen CM, Polvsen JV, Ingerslev J 1985 Application of an enzyme-linked immunosorbent assay (ELISA) for the measurement of small quantities of alpha-2-macroglobulin. Scan J Clin Lab Invest 45: 735-740.
Polvesen JV, Ingerslev J, Petersen CM 1987. Application of an enzyme-linked immunoassay for the measurement of pregnant zone protein (PZP) in cell culture supernatants and sera. Scand J Clin Lab Invest 7: 207-213.
Shibuya N, Berry JE, Goldstein IJ 1988. One-step purification of murine IgM and human alpha-2-macroglobulin by affinity chromatography on immobilized snowdrop bulb lectin. Arch Biochem Biophys 267: 676-680.
Sottrup-Jensen L 1989. Alpha-macroglobulins: structure, shape and mechanism of proteinase complex formation. J Biol Chem 264:11539-11542.
Van Leuven F 1982. Human alpha-2-macroglobulin: structure and function. Trends Biochem Sci 7: 185-187.
Van Leuven F, Cassiman J-J, Van Den Berghe H 1985. Separation of slow and fast form human alpha-2-macroglobulin by hydrophobic high performance liquid chromatography. Sc Tools 32: 41-43.
Van Leuven F, Marynen P, Cassiman J-J, Van Der Berghe H 1987. A mouse monoclonal antibody to human alpha-2- macroglobulin (A2M) cross-reacts with A2M from mouse: epitope mapping and characterization of the subunit structure of murine A2M. J Biochem 101: 1181-1189.
Virca GD, Travis J, Hall PK, Roberts RC 1978. Purification of human alpha-2-macroglobulin by chromatography on cibacron blue sepharose. Anal Biochem 89: 274-278.
Copyright 1994 Fundacao Oswaldo Cruz - FIOCRUZ The following images related to this document are available:Line drawing images[oc94041d.gif] [oc94041a.gif] [oc94041c.gif] [oc94041b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}