 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Comparative Chromatin Analysis of Trypanosoma congolenseWolfram Schlimme, Markus Burri, Bruno Betschart, Hermann Hecker (+)
Code Number: OC94048
Size of Files:
Text: 19K
Graphics: Photographs (Jpg) - 184K / Halftones (Gif) - 172K
Swiss Tropical Institute, Postfach CH-4002 Basel, Switzerland
The chromatin of Trypanosoma congolense was analyzed by electron microscopy. The chromatin is organized as nucleosome filaments but does not form a 30 nm fiber. There are five groups of histones, including a histone H1-like protein, which has a molecular weight within the range of the core histones, and is extremely hydrophilic. Weak histone-histone interaction, a typical feature of trypanosome chromatin, was found. These results are similar to those for T. cruzi and T. b. brucei, but differ significantly from those for higher eukaryotes. The results confirm the model of trypanosome chromatin, and support the theory of their early separation from the other eukaryotes during the evolution. T. congolensis is an excellent model for chromatin research on trypanosomes, because it is easy to cultivate and its chromatin has, a relatively high stability, compared to that of other trypanosomes.
Key words: Trypanosoma congolense - chromatin structure - histones - lower eukaryotes - procyclic culture forms
Trypanosomes, protozoan parasites of man and animals, have various nuclear features different from those of higher eukaryotes. No condensed chromosomes can be seen during nuclear division (Vickerman & Preston 1970), and the compaction of the chromatin in the nucleus is less pronounced as compared to the chromatin of rat liver nuclei (Hecker & Gander 1985). The histones of the trypanosomes differ significantly from those of higher eukaryotes in amino acid composition and sequence (Bender et al. 1992b, Toro et al. 1992, Hecker 1993). There are weaker interactions, histone-histone and histone-DNA, and this leads to a reduction in chromatin stability (Hecker et al. 1989, Burri et al. 1993).
The chromatin of Trypanosoma b. brucei procyclic and of T. cruzi epimastigote culture forms was shown to be organized in a nucleosome filament-like form, but did not form a 30 nm fiber. Histone H1 was thought to be absent in trypanosomes for a long time, mainly because of the poor condensation behaviour of the chromatin, and also because no protein with the characteristics of histone H1 of higher eukaryotes was found (Hecker et al. 1994). Toro and Galanti (1988) showed for T. cruzi and Schlimme et al. (1993) for T. b. brucei that histone H1-like proteins do exist, but that these proteins are different in several ways from histone H1 of higher eukaryotes. They are very hydrophilic and have a molecular weight of about 13'000 D instead of the 21'000 D of H1 of higher eukaryotes (Van Holde 1989), and the globular part appears to be quite small (Schlimme et al. 1993).
Most investigations on the chromatin of trypanosomes have been carried out on T. cruzi and T. b. brucei (Hecker et al. 1994). In the study described here, the chromatin of T. congolense, the casual agent of African bovine trypa nosomiasis, an important disease of cattle and domestic animals, was investigated.
MATERIALS AND METHODS
T. congolense STIB 802 procyclic culture forms were used. They were grown in SM medium containing 20% heat-inactivated fetal bovine serum, 10 ug/ml hemin, 10 ug/ml gentamicin, 5% CO2 at 27 oC (Cunningham 1977). T. cruzi Y strain epimastigotes were cultured in modified Mattei medium (Mattei et al. 1977) containing 10% heat-inactivated fetal bovine serum, 10 ug/ml hemin, 10 ug/ml gentamicin, 30 mM leucine, at 27 oC.
T. b. brucei STIB 345 AB strain procyclic culture forms were cultured as described by Brun and Schonenberger (1979).
Exponentially growing procyclic trypa nosomes (2 to 3x10^10) were harvested and nuclei were prepared as described for procyclic culture forms of T. b. brucei (Schlimme et al. 1993).
Preparation of soluble chromatin - Nuclei of T. congolense were digested with 0.2 units micrococcal nuclease (Sigma, N-3755) per 20 A260 at 30 oC for 50s. Nuclei of rat liver were digested with 0.4 units micrococcal nuclease per 20 A260 at 30 oC for 50s. The nuclei were pelleted, and the chromatin solubilized by nuclear lysis in a hypotonic buffer containing 1 mM triethanolaminehydrochloride (TEACI) and 0.2 mM Na2EDTA, pH 7.4. Insoluble material was removed by centrifugation (Thoma et al. 1979, Hecker & Gander 1985).
Gradient analysis of the chromatin digest - Gradient analysis was done in 5.5 - 28.5% (w/v) 17 ml isokinetic sucrose gradients containing 5 mM TEACI pH 7.4, 0.2 mM Na2EDTA and 10 mM NaCl (Noll 1969). Centrifugation was performed for 14 hr at 25'000 g in a Kontron TST 28/17 swing out rotor. The gradients were monitored at 254 nm and the bottom fractions containing the larger fragments of soluble chromatin were used for analysis by electron microscopy (Thoma & Koller 1981).
Electron microscopy - The fractions with the large chromatin fragments were divided into four aliquots and dialyzed against 5 mM TEACI (pH 7.4), 0.2 mM Na2EDTA containing 0, 10, 40 or 100 mM NaCl respectively for 4 hr. Then the dialysis buffer was made 0.1% (v/v) with glutaraldehyde and samples were fixed at least for 15 hr and prepared for EM observation (Thoma et al. 1979).
Histone extraction from purified nuclei - Nuclei were resuspended in 0.25 NHCl (Elpidina et al. 1979) or in 5% (v/v) perchloric acid (PCA) (Sanders 1977) and proteins extracted for 60 min under constant agitation. Insoluble material was pelleted at 4000 g for 5 min. The supernatants, containing the histones, were removed and dialyzed against 1 mM TEACI, 0.2 mM Na2EDTA pH 7.4 and lyophilized. All the preparations were carried out at 0-4 oC if not otherwise stated.
SDS Tricine PAGE (Schagger & von Jagow 1987) - Lyophilized histones were solubilized in sample buffer, and separated in a linear 17.6% SDS Tricine polyacrylamide gel as described (Schlimme et al. 1993).
Triton acid urea polyacrylamide gel electrophoresis (Alfageme et al. 1974) - Lyophilized histones were dissolved in sample buffer, and electrophoresis in a linear 15% triton DF-16 acid urea polyacrylamide gel was carried out as described (Schlimme et al. 1993).
RESULTS
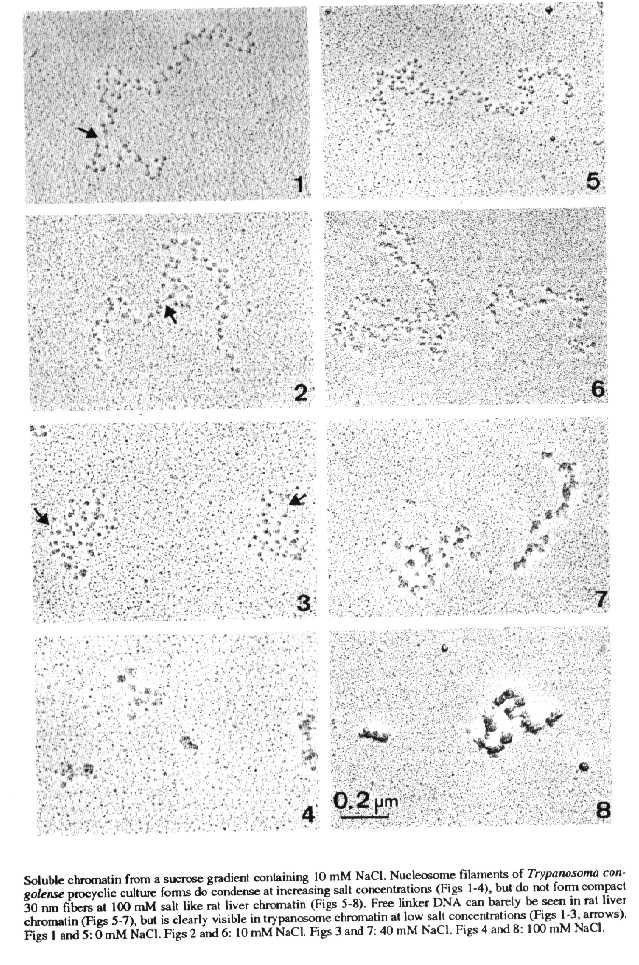
Structure and compaction pattern of soluble chromatin - Soluble chormatin of T. congolense, centrifuged through a sucrose gradient containing 10 mM NaCl, dialyzed against concentrations of 0, 10, 40, or 100 mM NaCl, and prepared for EM observation, is organized in the form of nucleosome filaments. The soluble chromatin shows some condensation at increasing ionic strength (Figs 1-4). However, solenoids (30 nm fibers) which are typical for rat liver chromatin (Figs 5-8), are not formed. Free linker DNA can barely be seen in rat liver chromatin (Figs 5-7), but is clearly visible in trypanosome chromatin at low salt concentrations (Figs 1-3).
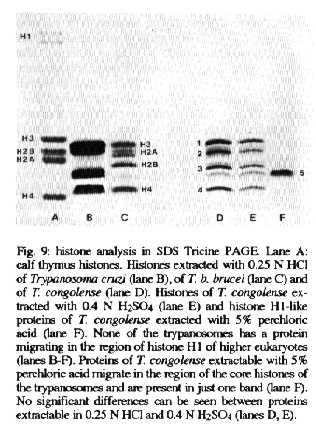
SDS Tricine PAGE - Histones of higher eukaryotes separate into four core histones and into two H1 variants in SDS Tricine PAGE (Fig. 9, lane A). Core histones of trypanosomes and higer eukaryotes differ in their electrophoretic mobility and number of variants and/or modifications. The trypanosome histones H3 and H4 have a very similar migration behaviour, while histones H2A and H2B are quite heterogeneous (Fig. 9, lanes B-E). Histone 2 of T. congolense has three variations, and histone 3 has two (Fig. 9, lane D, E). Neither T. cruzi, T. congolense nor T. b. brucei has any protein migrating in the region of histone H1 of higher eukaryotes (Fig. 9, lanes B-E). Proteins extracted with 5% PCA migrate in the lower region of the core histones and are metachromatically stained with Coomassie brilliant blue (Fig. 9, lane F). These PCA-extractable proteins of T. congolense migrate in only one band.
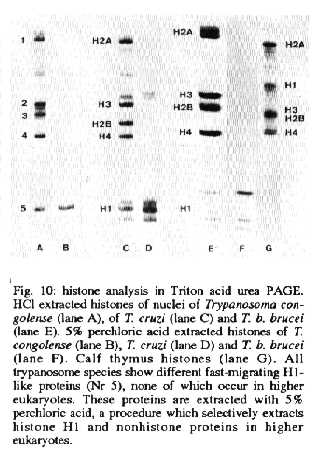
Triton acid urea PAGE - In Triton acid urea gels, which separate proteins according to their hydrophobicity, the core histones of T. congolense migrate in four main complexes 1, 2, 3 and 4 (Fig. 10, lane A). Note that the numerical sequences of the histones in the SDS and Triton gel systems do not correspond. A fast migrating, very hydrophilic complex of H1-like proteins (Nr. 5), (Fig. 10, lanes A-F). can be seen. No proteins of higher eukaryotes can be seen in the region of H1-like proteins (lane G). Although the protein complexes 1-4 do run in the same region as the core histones of higher eukaryotes (Fig. 10, lane G), all the histones of the trypanosomes differ from those of higher eukaryotes.
Differences exist between the histones of various trypanosomes species in the number of bands, their position in the gel and their relative amount. This is specially true for the H1-like proteins (Fig. 10, lanes A-F, complex Nr. 5). All the proteins in this area can be selectively extracted with 5% perchloric acid. These proteins are metachromatically stained with Coomassie brilliant blue. The H1-like proteins can also selectively be removed by 500 mM NaCl (not shown).
DISCUSSION
The banding pattern of the core histones of T. congolense in Triton acid urea gels differs significantly from that of histones from higher eukaryotes and also from that of other trypanosomes. This indicates that the core histones have different amino acid compositions and sequences. In SDS Tricine gels the core histones of trypanosomes migrate in the same region as those of higher eukaryotes, which means that they have a similar molecular weight. Histones are known to be highly conserved proteins. Among them histone H4 is the most conserved, followed by H3, H2A, H2B and H1 (Van Holde 1989). Bander et al. (1992b) and Toro et al. (1992) showed for T. b. brucei and T. cruzi a sequence divergence in trypanosomes of about 35% in the N- and C-terminal regions even for histone H4, suggesting a very early separation of the trypanosomes from other eukaryotes during evolution (Hecker 1993). The observation that the band corresponding to H4 in T. congolense ran in the same position in both gel systems as H4 of the other trypanosomes further supports this theory. The positions in the SDS gel of the proteins 2 and 3, the counterparts of histone H2A and H2B (Bender et al. 1992a, b) are quite variable between the different trypanosome subgroups, and the considerable differences between the histones H2A and H2B reflect this fact.
In T. congolense, a histone H1-like protein is present, but does not have as many variants or modifications as its counterparts in T. cruzi or T. b. brucei. Like H1 of the other trypanosomes, it is extractable with 5% PCA and shows meta-chromasia, which is a sign of a high content of lysine (Duhamel et al. 1980). Furthermore it can be selectively removed by 500 mM NaCl. The similarity of the migration behaviour of this protein as compared to other trypanosome H1-like proteins Duschak and Cazzulo (1990), and all the other properties mentioned above, allow the conclusion that the PCA-extractable proteins of T. congolense correspond to the H1-like proteins of other trypanosomes. However, the question how such a small histone H1 can execute its role in chromatin condensation is not yet answered. The poor condensation of the chromatin of T. congolense might be due to the small size of this H1-like protein. Genetically inactive chromatin of higher eukaryotes is organized as nucleosome filaments and forms 30 nm fibers (solenoids) at 100 mM NaCl (Thoma et al. 1979). The chromatin of T. congolense is also organized in the form of nucleosome filaments, but does not form 30 nm fibers. The compaction pattern of the chromatin of T. congolense procyclic culture forms is comparable to that of T. b. brucei blood stream forms (Schlimme et al. 1993) and to that of T. cruzi epimastigote culture forms (Hecker et al. 1994), but is more pronounced than that of T. b. brucei procyclic culture forms (Hecker & Gander 1985, Schlimme et al. 1993). Trypanosomes do not form chromosomes sensu stricto, and we do not know, whether a high degree of condensation is necessary for the function of the genetic system, as it is in higher eukaryotes. One of the main questions that still remains is what type of condensation in trypanosome chromatin is achieved. The stability of chromatin depends on the nature and strength of the protein-protein and the protein-DNA interactions (Yager et al. 1989). The stability of the soluble chromatin of T. congolense is reduced as compared to that of higher eukaryotes. The interactions of histones and DNA are weak and easily destabilized by experimental conditions, as described for other trypanosomes (Hecker et al. 1989, Bender et al. 1992c, Schlimme et al. 1993, Burri et al. 1993).
Differences in the stability of the chromatin among the trypanosomes exist not only between the different species (Hecker et al. 1994), but also between different stages of the life cycles (Schlimme et al. 1993).
The chromatin of T. congolense is a very good model for chromatin research in trypanosomes, since it has some major advantages as compared to the other trypanosomes investigated so far. First, T. congolense procyclic culture forms are easy to cultivate in vitro in large numbers. Second, the instability of the soluble chromatin is not as pronounced as in T. b. brucei procyclic culture forms. Therefore reconstitution of soluble chromatin and other experiments to investigate the condensation behaviour can be better carried out with this parasite. Last but not least, makes the fact, that T. congolense is not pathogenic for humans, it is very convenient to work with.
ACKNOWLEDGEMENTS
To Jennifer Jenkins for critical discussion of the manuscript.
REFERENCES
Alfageme CR, Zweidler A, Mahowald A, Cohen LH 1974. Histones of Drosophila Embryos. J Biol Chem 249: 3729-3736.
Bender K, Betschart B, Schaller J, Kampfer U, Hecker H 1992a. Biochemical properties of histone-like proteins of procyclic Trypanosoma brucei brucei. Acta Trop 50: 169-184.
Bender K, Betschart B, Schaller J, Kampfer U, Hecker H 1992b. Sequence differences between histones of procyclic Trypanosoma brucei brucei and higher eukaryotes. Parasitology 105: 97-104.
Bender K, Betschart B, Hecker H 1992c. Histone-DNA interactions in the chromatin of procyclic Trypanosoma brucei brucei. Parasitol Res 87: 495-500.
Brun R, Schonemberger M 1979. Cultivation and in vitro cloning of Tripanosoma brucei in a semi-defined medium. Acta Trop 36: 289- 292.
Burri M, Schlimme W, Betschart B, Kampfer U, Schaller J, Hecker H 1993. Biochemical and functional characterization of histone H1- like proteins in procyclic Trypanosoma brucei brucei. Parasitol Res 79: 649-659.
Cunningham I 1977. New culture medium for maintenance of tsetse tissues and growth of trypanosomatids. J Protozool 24: 325-329.
Duhamel RC, Meezan E, Brendel K 1980. Metachromatic staining with Coomassie-Brilliant Blue R-250 of the proline-rich calf thymus histone H1. Biochim Biophys Acta 626: 432-442.
Duschak VG, Cazzulo JJ 1990. The histones of the insect trypanosomatid, Crithidia fasciculata. Biochim Biophys Acta 1040: 159-166.
Elpidina EN, Zaitseva GN, Krasheninnikov JA 1979. Histones from Trypanosoma lewisi nuclei. Biokhimiya 44: 1830-1841.
Hecker H 1993. Man and sea urchin - More closely related than African and American trypanosomes? Parasitol Today 9: 57.
Hecker H, Gander ES 1985. The compaction pattern of the chromatin of trypanosomes. Biol Cell 53: 199-208.
Hecker H, Bender K, Betschart B, Modespacher UP 1989. Instability of the nuclear chromatin of procyclic Trypanosoma brucei brucei. Mol Biochem Parasitol 37: 225-234.
Hecker H, Betschart B, Bender K, Burri M, Schlimme W 1994. The chromatin of trypanosomes. Int J Parasitol (in press).
Mattei DM, Goldberg S, Morel CM, Azevedo HP, Roitman I 1977. Biochemical strain characterization of Trypanosoma cruzi by restriction endonuclease cleavage of kinetoplast DNA. Fed Europ Biochem Soc Let 74: 264-268.
Noll H 1969. An automatic high-resolution gradient analyzing system. Anal Biochem 27: 130-149.
Sanders C 1977. A method for the fractionation of the high- mobility group non-histone proteins. Biochem Biophys Res Comm 78: 1034-1042.
Schagger H, von Jagow G 1987. Tricien-sodium-dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal Biochem 166: 368-379.
Thoma F, Koller T 1981. Unravelled nucleosomes, nucleosome beads and higher order structures of chromatin: Influence of non-histone components and histone H1. J Mol Biol 149: 709-733.
Thoma F, Koller T, Klug A 1979. Involvement of histone H1 in the organization of the nucleosome and the salt-dependent superstructures of chromatin. J Cell Biol 83: 403-424.
Toro CG, Galanti N 1988. H1 histone and histone variants in Trypanosoma cruzi. Exp Cell Res 174: 16-24.
Toro CG, Wernstedt C, Medina C, Jaramillo N, Hellman U, Galanti N 1992. Extremely divergent Histone-H4 sequence from Trypanosoma cruzi - evolutionary implications. J Cell Biochem 49: 266-271.
Van Holde KE 1989. Chromatin. Springer Series in Molecular Biology. New York: Springer Verlag, 497 pp.
Vickerman K, Preston TM 1970. Spindle microtubules in the dividing nuclei of trypanosomes. J Cell Sci 6: 365-383.
Yager TD, Mc Murray T, van Holde KE 1989. Salt-induced release of DNA from nucleosome core particles. Biochemistry 28: 2271-2281.
Copyright 1994 Fundacao Oswaldo Cruz - FIOCRUZ The following images related to this document are available:Halftone images[oc94048a.gif] [oc94048b.gif] [oc94048c.gif]Photo images[oc94048a.jpg] [oc94048b.jpg] [oc94048c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}