 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Extramedullary Hematopoiesis in Murine Schistosomiasis Mansoni HL Lenzi+, JA Lenzi, FC Rosman, M Pelajo-Machado, EM Mota, MS Panasco, DN Oliveira Departamento de Patologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
Code Number: OC95036
Size of Files:
Text: 17K
Graphics: Line Drawings (gif) - 24K
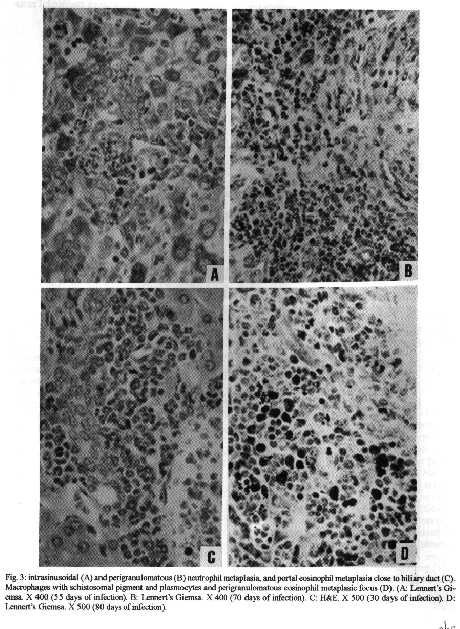
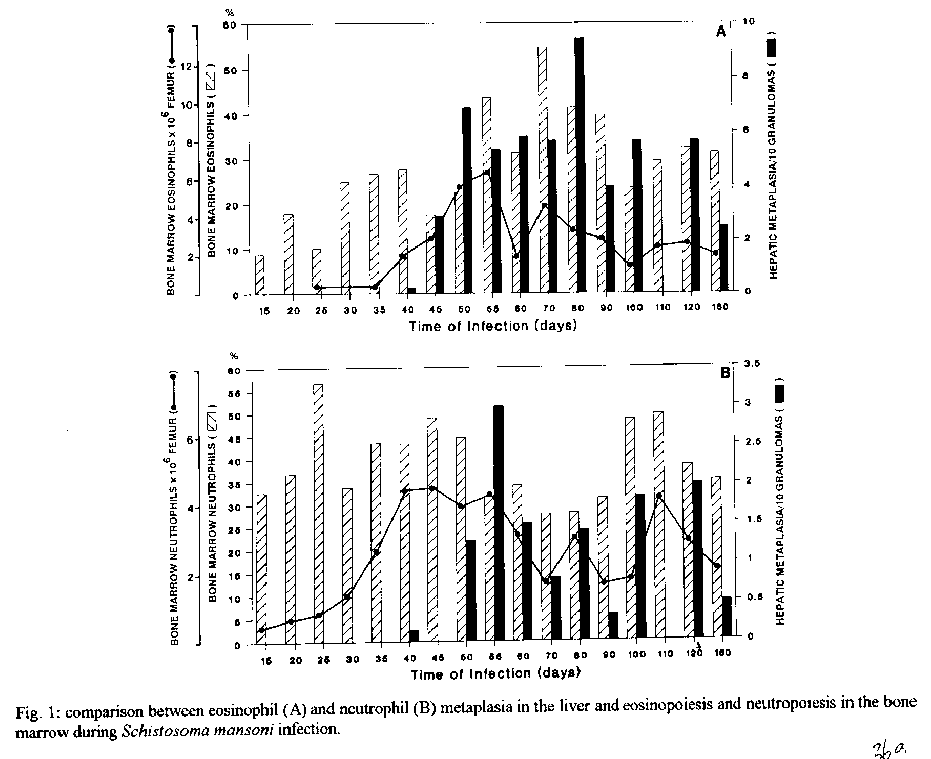
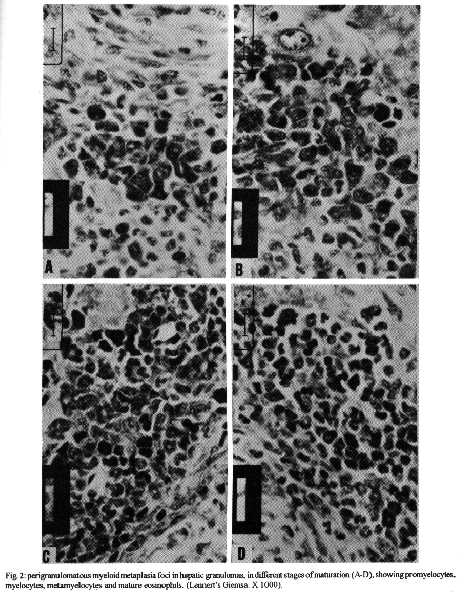
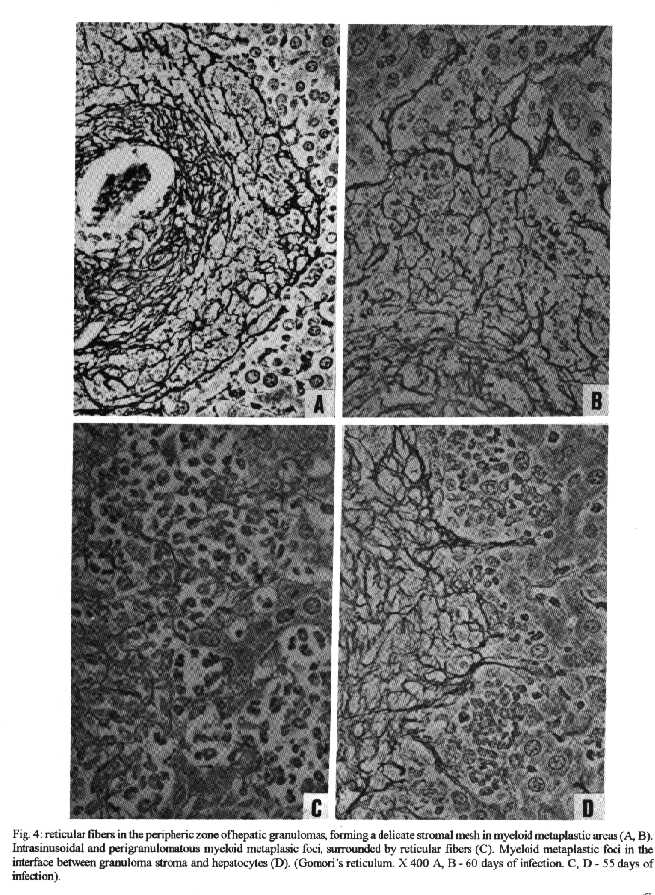
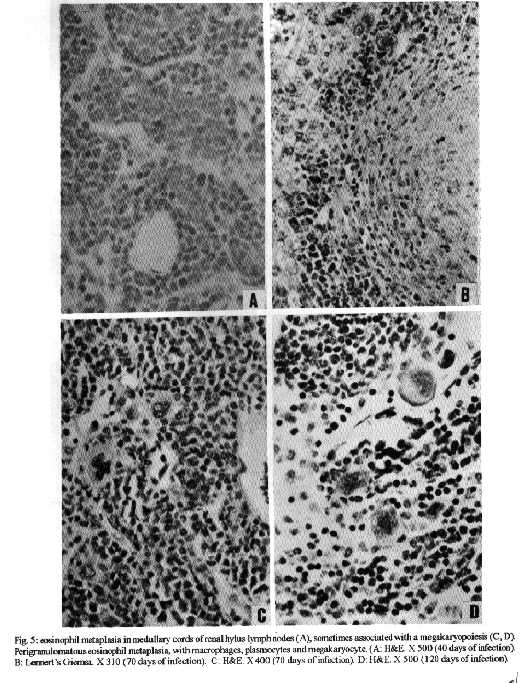
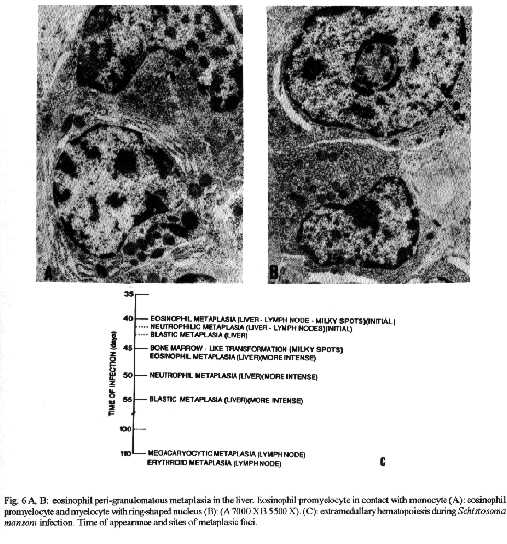
Photos (jpg) - 350K / Halftones (gif) - 254K'During Schistosoma mansoni infection, there is morphological evidence of involvement of various hematopoietic growth factors, which cause eosinophil, neutrophil, megakaryocytic and erythroid extramedullary foci in the liver, lymph nodes and omental and mesenteric milky spots. While the eosinophil metaplasia in the periphery of hepatic granulomas roughly reproduced the intensity of the medullary eosinopoiesis, the neutrophil metaplasia, on the contrary, was more intense during the period of neutrophil depression in the bone marrow. This fact suggests that extramedullary hematopoietic foci are locally regulated, and amplify and/or compensate the systemic hematopoietic response during the infection. Key words : eosinophil - neutrophil - megakaryocyte - hematopoiesis - Schistosoma mansoni - lymph nodes - milky spots Schistosoma mansoni infection in mice and humans, due to deposition of eggs in host tissues and systemic and local release of antigens, causes a very complex sequence of events, changing the behavior of eosinophil, neutrophil, monocytic, megakaryocytic, lymphocytic and erythroid series inside the bone marrow (Lenzi et al. 1987, Lenzi 1991). However, in mice and hamsters infected with S. mansoni or S. japonicum, extramedullary islands of eosinophil precursors regularly occur in the liver, colonic mesenteries, and mesenteric lymph nodes (Byram et al. 1978, Borojevic et al. 1981). In this study we showed the time of occurence and the different places where extramedullary hematopoiesis were detected during schistosome infections. We also related the intensity of hepatic peri-granulomatous myeloid metaplasia with the medullary myelopoiesis. Swiss Webster mice of both sexes were studied on days 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 70, 80, 90, 100, 110, 120 and 160 after infection. The animals were infected when they were five days old by percutaneous exposure to 70 cercariae of the Belo Horizonte strain of S. mansoni. Six animals were sacrificed each day, together with the same number of matched controls. Femoral bone marrow cells were isolated by a modification of the method described by van Furth and Cohn (1968), cytocentrifuged and stained with Giemsa-Glycine (Lenzi 1991). Absolute eosinophil and total nucleated cells counts were carried out using Discomb's fluid in Fuchs-Rosenthal and Turk's fluid in Neubauer chambers, respectively. The total number of medullary neutrophils was figured out based on the percentage of each lineage in the cytocentrifuged slides, and the total cell counting in the Neubauer chamber. Samples were taken from the liver, intestines, pancreas, lungs, spleen, lymph nodes and mesentery, fixed in Millonig formalin, and embedded in paraffin. Sections were stained with hematoxilin and eosin, Lennert's Giemsa and Gomori's silver reticulin stains. To quantify the number of hepatic granulomas presenting myeloid metaplasia foci, ten of them, stained with Lennert s Giemsa, were studied. For electron microscopic examination, liver sections were fixed in 2.5 % glutaraldeyde buffered with 0,1 M cacodylate and postfixed in 1% osmium tetroxide, dehydrated in graded acetone series and embedded in epoxy resin. Ultra-thin sections were stained with uranyl acetate and lead citrate and observed by an EMS 10 B Zeiss electron microscope. The most precocious finding was an increase in splenic eosinopoiesis, erythropoiesis, megakaryopoiesis and neutropoiesis that was verified on the 25th day of infection. Foci of extragranulomatous metaplasia characterized by immature myeloid cells in portal space (Fig. 3-C), sinusoids (Fig. 3-A) and close to the capsule were detected between 30 and 35 days after infection. Extramedullary eosinophilic granulocytopoiesis occurred in the periphery of hepatic schistosomal granulomas from the 40th day on, attaining maximum intensity on the 80th day after infection, persisting until the end of the experiment (Fig. 1-A). Peri-granulomatous neutrophil metaplasia also appeared from the 40th day onward, but it was more intense on the 55th day of infection. While the eosinophil metaplasia roughly reproduced the intensity of the medullary eosinopoiesis, the neutrophil metaplasia, on the contrary, was more intense during the period of neutrophil depression in the bone marrow (Fig. 1-B). The myeloid foci were composed by promyelocytes, myelocytes, metamyelocytes and more mature eosinophil or neutrophil granulocytes (Figs 2, 3, 6-A,B). The myeloid foci or nests were localized in the periphery of the granulomas (external zone) and the cells were in direct contact with hepatocytes and intermixed with or surrounded by reticular and type III collagen fibers (Fig. 4). With the advancing of the infection, the metaplasic foci exhibited a predominance of mature cells over blast cells. Macrophages rich in schistosome pigment were often found in the middle of perigranulomatous cellular metaplasia (Fig. 3- D). Omental and mesenteric milky spots and lymph nodes mainly from mesentery and renal hilus presented, very often, extramedullary hematopoiesis, expressing eosinophil, and, eventually, neutrophil, megakariocytic and erythroid cells (Figs 5, 6-C). Myeloid metaplasia was not found in control mice and in the intestinal and pancreatic granulomas of the infected group. Only in a single case, with 160 days of infection, one pulmonary granuloma showed eosinophil metaplasia. The organ dependency can be due to local capacity to produce hemopoietic growth factors and reproduce hemopoietic inductive microenvironment (Jenkins et al. 1972). Though a wide variety of murine cell types have been shown to produce specific hemopoietic growth factor mRNAs and bioactivities in vitro (Nicola 1989), little is known about the sites of their production in vivo (Troutt & Lee 1989). Granulocyte-macrophage (GM) and Granulocyte (G) colony stimulating factors (CSF) activities have been partially characterized in a variety of mouse tissues. However, definitive evidence at the molecular level for their in vivo production in these tissues has been lacking (Troutt & Lee 1989). For example, Chan et al. (1986) have shown no evidence of GM-CSF mRNA production in any of 57 human tissue samples tested. In contrast, M-CSF mRNA has been found in all murine tissues tested except for intestine (Rajavashisth et al. 1987). This fact can explain the lack of hemopoietic foci in intestinal granulomas, suggesting that induced transcription of hemopoietic growth factor genes is involved in vivo in the host response to infection, and that the production of some of these factors may be compartmentalized in the organism (Troutt & Lee 1989). Probably, during schistosomal infection there is supply of bone marrow-derived cells to various tissues. The finding of immature myeloid cells within the hepatic sinusoids (Fig. 3-A) before the occurrence of perigranulomatous eosinophil metaplasia, suggests the exogenous origin of the eosinophil precursors, as was also observed by Borojevic et al. (1981). Clark et al. (1988) proposed that granuloma macrophages are supplied by both bone marrow-derived and focally produced monocytes in S. mansoni-infected mice. The relative contributions of these two sources to the monocyte supply are subject to change as the vigorous granulomatous response undergoes immunologic down modulation. It has been shown that cells capable of division and differentiation do circulate normally in peripheral mice blood. Goodman and Hodgson (1962) showed clearly that normal peripheral leukocytes from F1 hybrid mice can be transplanted and proliferate in lethally irradiated parent or closely related homologous strain animals to promote survival and to give rise to lymphopoiesis, granulopoiesis and erythropoiesis of donnor type. Butterfield et al. (1982) demonstrated that eosinophil colonies developed mainly from nonrosetting, null cell population. Eosinophil-Co- lony Forming Cell (Eo-CFC) lacked the human T-cell marker Leu 1, but demonstrated the human B-cell marker, HLA-DR. Eo-CFC was also present in peripheral blood and possessed properties similar to those in the bone marrow. These fact may explain the hemopoietic cells seeding in tissues of hemopoietic potential, such as lymph nodes, milky spots and liver granulomas. The selective stimulation of eosinophils, neutrophils and, maybe, monocytic series in the periphery of hepatic granulomas could be achieved either by progenitor selection or through specific stimulation of less differentiated stem cells (Borojevic et al. 1981). Schistosomal granulomas are composed by cells that are potential producers of hemopoietic growth factors, such as lymphocytes, macrophages, fibroblasts, mast cells and endothelial cells ( Broudy et al. 1986, Clark et al. 1988, Zsebo et al. 1988, Nicola 1989). Borojevic et al. (1981) considered the Kupffer cell stimulation and hiperplasia responsible for this extramedullary granulocytopoiesis. Borojevic et al. (1985) observed that intraperitoneal glass implants in mice with chronic S. mansoni infection induce intense local myeloid reactions involving essentially myeloid granulocytes. An association of myelopoiesis with differentiated macrophages mobilized on glass implants and with dense ameboid cells located inside myeloid foci was detected. A macrophage-dependent induction of this eosinophil reaction was postulated. An augmentation of the mutual interaction of T cells and macrophages by GM-CSF is one of the important mechanisms of eosinophil-rich granuloma formation in S. japonicum (Owhashi et al. 1987). The simultaneous occurrence of hemopoietic foci in different organs suggests that, during murine schistosomal infection, stimulating common factors are acting systematically in various sites at the same time, or they are locally produced by similar stimulants. Actually, the levels of bone-marrow and perigranulomatous eosinopoiesis showed here followed the same kinetics of splenic lymphocyte interleukin-5 response (Th2 dependent) described by Grzych et al. (1991) in the same infection. Homing of progenitor cells can be conceived as a two-step phenomenon: first, progenitor cells arriving from bone marrow must be recognized by and interact with the luminal surface of endothelium; second, the progenitor cells must be recognized by and bind to lineage-specific stromal cells that support their proliferation and maturation. Extracellular matrix components can take part in this phenomenon. For instance, it has been shown that one class of proteoglycans, heparan sulfate, can selectively extract and bind hematopoietic growth factors, presumably presenting them to progenitor cells (Roberts et al. 1988). Another fairly well-defined protein called haemonectin has been shown to be involved in the lineage specific binding of granulocytic precursors during both postnatal and fetal development (Campbell et al. 1987). Junqueira et al. (1986) demonstrated that glycosaminoglycan content was increased in schistosome granulomas, and we observed a topographical relationship between the connective tissue and the perigranulomatous eosinophil foci (Fig. 4). Our findings indicate that, during schistosome infection, there is morphological evidence of involvement of various hematopoietic growth factors, which act in different sites, specially around the hepatic granulomas, and in lymph nodes and milky spots, causing extramedullary hematopoiesis. The hematopoietic foci appeared to be locally regula-ted, and amplify and/or compensate the systemic or bone marrow hematopoietic response caused by a huge antigen release by the parasites during the infection. ACKNOWLEDGMENTS To AS Rodrigues, FF Cruz, H Ferreira, ID Pedro, LFG Caputo and VC Valentin for technical assistance and to HMN Diniz, GJ Vieira, JC Cruz and VCR S for preparing the figures. REFERENCES Borojevic R, Nicola MH, Santos-da-Silva C, Grimaldi Jr G 1985. Schistosoma mansoni: extramedullar eosinophil myelopoiesis induced by intraperitoneal glass implants in chronically infected mice. Exp Parasitol 59: 290-299. Borojevic R, Stocker S, Grimaud JA 1981. Hepatic eosinophil granulocytopoiesis in murine experimental schistossomiasis mansoni. Br J Exp Pathol 62: 480-489. Broudy VC, Kaushansky K, Segal GM, Harlan JM, Adamson JW 1986. Tumor necrosis factor type alfa stimulates human endothelial cells to produce granulocyte/macrophage colony-stimulating factor. Proc Natl Acad Sci USA 83: 7467- 7471. Butterfield JH, Eisenbrey AB, Gleich GJ 1982. Membrane marker characterization of the eosinophil colony-forming cell. Br J Haematol 51: 209-216. Byram JE, Imohiosen EAE, von Lichtenberg F 1978. Tissue eosinophil proliferation and maturation in Schistosome- infected mice and hamsters. Am J Trop Med Hyg 27: 267-270. Campbell AD, Long MW, Wicha MS 1987. Haemonectin, a bone marrow adhesion protein specific for cells of granulocyte lineage. Nature 329: 744-746. Chan JY, Slamon DJ, Nimer SD, Golde DW, Gasson JC 1986. Regulation of expression of human granulocyte/macrofage colony-stimulating factor. Proc Natl Acad Sci USA 83: 8669-8673. Clark CR, Chen BDM, Boros DL 1988. Macrophage progenitor cell and colony-stimulating factor production during granulomatous schistosomiasis mansoni in mice. Inf Immunity 56: 2680-2685. Goodman JW, Hodgson GS 1962. Evidence for stem cells in the peripheral blood of mice. Blood 19: 702-714. Grzych JW, Pearce E, Cheever A, Caulada ZA, Caspar P, Heiny S, Lewis F, Sher A 1991. Egg deposition is the major stimulus for the production of Th2 cytokines in murine schistosomiasis mansoni. J Immunol 146: 1322-1327. Jenkins VK, Trentin JJ, Speirs RS, McGarry MP 1972. Hematopoietic Colony Studies. VI. Increased eosinophil- containing colonies obtained by antigen pre-treatment of irradiated mice reconstituted with bone marrow cells. J Cell Physiol 79: 413-422. Junqueira LCU, Montes GS, Toledo OMS, Joazeiro PP 1986. Morphological, histochemical and biochemical observations on the connective tissue matrix of in situ and isolated hepatic granulomas in experimental murine schistosomiasis. Ann Trop Med Parasitol 80: 27-41. Lenzi HL 1991. A dinƒmica da resposta hematol¢gica e celular na esquistossomose mansonica murina, com enfase nas series eosinof¡lica e mastocit ria. Doctoral Thesis. Universidade Federal de Minas Gerais, Brazil, 580 pp. Lenzi HL, Sobral ACL, Lenzi JA 1987. "In vivo" kinetics of eosinophils and mast cells in experimental murine schistosomiasis. Mem Inst Oswaldo Cruz 82 (Suppl.IV): 67-76. Nicola NA 1989. Hemopoietic cell growth factors and their receptors. Annu Rev Biochem 58: 45-47. Owhashi M, Maruyama H, Nawa Y 1987. Granulocyte-macrophage colony-stimulating factor enhances the production of eosinophil chemotactic lymphokine by egg-associated granulomas of Schistosoma japonicum-infected mice. Infect Immun 55: 2042-2046. Rajavashisth TB, Eng R, Shadduck RK, Waheed A, Ben-Avram CM, Shively JE, Lusis AJ 1987. Cloning and tissue-specific expression of mouse macrophage colony-stimulating factor mRNA. Proc Natl Acad Sci USA Biochem 84: 1157-1161. Roberts R, Gallagher J, Spooncer E, Allen TD, Bloomfield F, Dexter TM 1988. Heparan sulphate bound growth factors: a mechanism for stromal cell mediated haemopoiesis. Nature 332: 376-378. Troutt AB, Lee F 1989. Tissue distribution of murine hemopoietic growth factor mRNA production. J Cell Physiol 138: 38-44. Van Furth R, Cohn ZA 1968. The origin and kinetics of mononuclear phagocytes. J Exp Med 128: 415-433. Zsebo KM, Yuschenkoff VN, Schiffer S, Chang D, McCall E, Dinarello CA, Brown MA, Altrock BC, Bagby Jr GC 1988. Vascular endothelial cells and granulopoiesis: Interleukin-1 estimulates release of G-CSF and GM-CSF. Blood 71: 99-103. Copyright 1995 Fundacao Oswaldo Cruz (Fiocruz)
The following images related to this document are available:Halftone images[oc95036b.gif] [oc95036f.gif] [oc95036d.gif] [oc95036e.gif]Photo images[oc95036d.jpg] [oc95036f.jpg] [oc95036c.jpg] [oc95036e.jpg] [oc95036b.jpg]Line drawing images[oc95036a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}