 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz, Vol.90(2):179-184 mar./apl. 1995 Molecular Aspects of Schistosoma mansoni Female Maturation Ana Lucia Moraes Giannini, Sergio Vasconcelos Linhares, Elena Cristina Caride, Vania Maria Martins Braga, Franklin David Rumjanek^+ + Corresponding author Departamento de Bioquimica Medica, ICB/CCS, Universidade Federal do Rio de Janeiro, Cidade Universitaria, Ilha do Fund o, Caixa Postal 68041, 21941-590 Rio de Janeiro, RJ, Brasil This work received financila sipport from the UDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases, CNPq, CAPES, FINEP and the Rockefellar Foundation.
Code Number: OC95037
Size of Files:
Text: 18K
Graphics: Line Drawings (gif) - 27K
Photos (jpg) -220K / Halftones (gif) - 341K
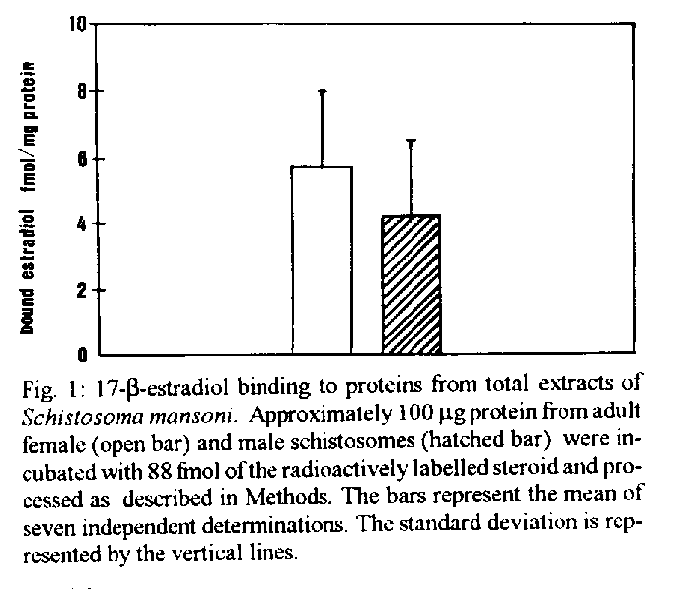
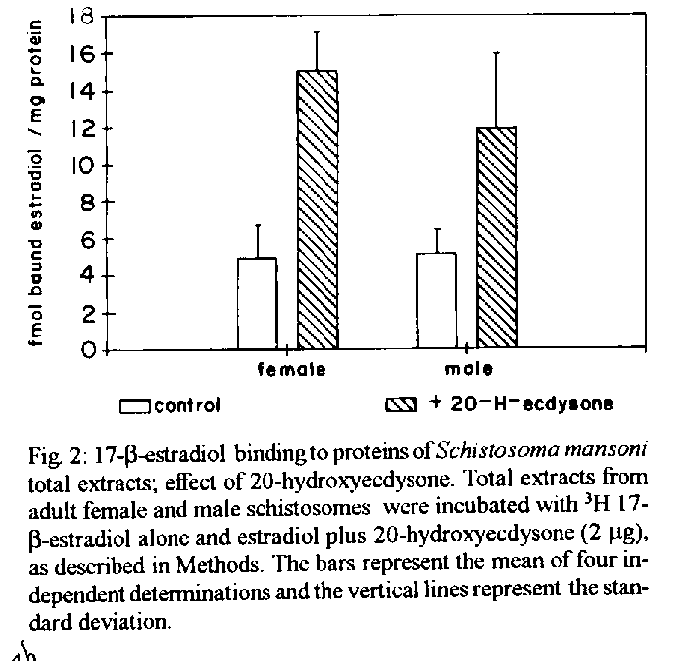
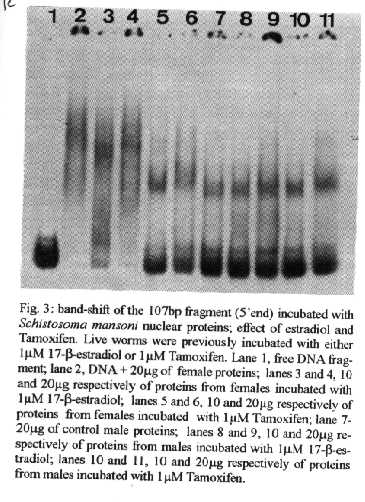
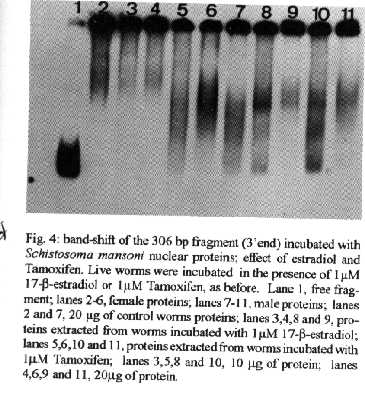
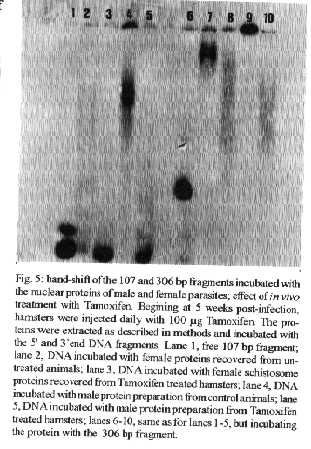
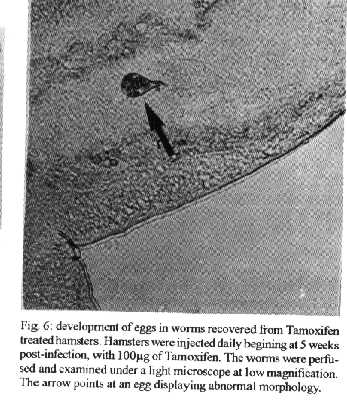
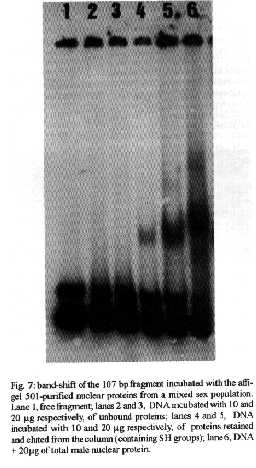
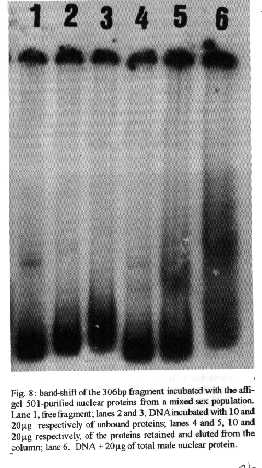
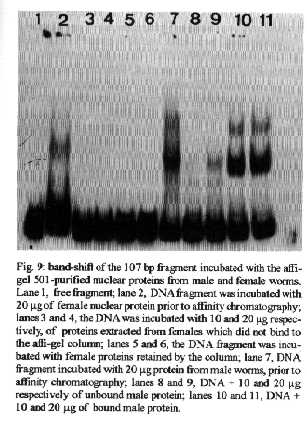
[TABLE AT END OF TEXT]Incubation of total protein extracts of Schistosoma mansoni with ^3H 17-b-estradiol and 20-hydroxyecdysone, revealed steroid binding proteins in both, male and female worms. The interaction of nuclear proteins with restriction fragments of the gender and stage-specific gene F-10 was investigated using the "Band-Shift" technique. Distinct male and female nuclear proteins bound to the fragments of this gene. Among the nuclear proteins, only those rich in cysteine residues bound to DNA. In vitro incubation of live worms with the estrogen antagonist Tamoxifen, altered the pattern of the DNA binding proteins, producing in females, a band profile similar to that obtained with male worm protein extracts. When Tamoxifen was injected into schistosome infected mice, the eggs produced by females presented an abnormal morphology, compatible with non-viable eggs. These results suggest that the regulation of transcription of the F- 10 gene might involve steroid receptors. Key words: Schistosoma mansoni - nuclear proteins - F- 10 gene - steroid receptors - Tamoxifen - eggs Based on the detection of consensus sequences present in stage and gender-specific genes of several species of schistosomes, the participation of regulatory proteins has been postulated (Bobek et al. 1989, 1991, Henkle et al. 1990, Chen et al. 1992). The downstream, non-translated region of the Schistosoma mansoni gene F-10 (Simpson et al. 1987, Kunz et al. 1987), is one such example, displaying a hexanucleotide and a pentanucleotide sequence, identical to hormone regulatory elements (HRE) present in genes regulated by steroids (Rumjanek et al. 1989). Preliminary attempts to characterize S. mansoni F-10 binding proteins, showed that nuclear proteins bound to different restriction fragments of the F-10 gene (Engelender et al. 1993). The occurrence of steroids in S.mansoni (Nirde et al. 1983) supports the idea that among the DNA binding proteins steroid receptors could be present. In the present work we have pursued this idea by carrying out experiments trying to verify: (a) whether estradiol and/or ecdysteroid binding proteins could be detected among the schistosome proteins; (b) whether the DNA binding proteins of S.mansoni shared some of the structural features of steroid receptors; and (c) whether Tamoxifen, an estrogen antagonist could affect the development of eggs. MATERIALS AND METHODS Parasites - Adult S.mansoni worms were obtained by perfusion of Syrian hamsters using the methods described by Smithers and Terry (1965). The worms were separated by sex using a fine brush and were either used immediately or stored at -70 C. Protein extracts were obtained according to methods described elsewhere (Goodwin 1990) in the presence of protease inhibtors. Protein extracts were stored at -70 C. Protein concentration was estimated using the Lowry method (Lowry et al. 1951). Nuclear proteins rich in thiol groups were purified by affinity chromatography using AFFI-GEL 501 (BioRad) according to the manufacturer's recommendations. Estradiol and Ecdysone Binding Assay - For the binding experiments, adult worms were homogenized in a Potter homogenizer in 10 mM Tris-HCl, pH 7, 7.5 mM MgCl2 at 4 C. The homogenate was centrifuged at 10,000 g for 3 min at 4 C and the supernatant collected, aliquoted and stored at -70 C until used. The binding assay consisted of incubating 100 ml of the protein extract in 10 mM Tris buffer pH 7.7 containing 0.25 M sucrose and 1 mM EDTA, with 90 fmol of 17-b-estradiol (2,4,6,7 ^3H-estradiol, 3.1-4.1 TBq/mmol) (Amersham), for 16hr at 4 C. The unbound ligand was removed by adding 0.3 mg of dextran coated charcoal for 10 min followed by centrifugation at 2500 RPM/ 10 min in an IEC bench-top clinical centrifuge. The supernatant was collected and counted in a liquid scintillation spectrometer. Controls for non-specific binding consisted of samples pre-incubated for 45 min with 20 ug of unlabeled 17-b-estradiol at 4 C. The ^3H-ligand was then added and incubated as above. Competition assays using 20- hydroxiecdysone (Sigma) were carried out as described above, except that 2 ug of the ecdysteroid were added to the incubation mixture. The F-10 Gene - The gene cloned in the pTZ plasmid vector (GeneScribe-Z), was obtained after digestion with EcoRI (Boehringer Mannheim) and purificatied from an agarose gel after eletrophoresis, using the QUIAEX kit (Quiagen). Pure F- 10 was digested with TaqI restriction enzyme (Pharmacia) and the fragments generated were recovered using the QUIAEX kit. The fragments were dephosphorylated with calf intestinal alkaline phosphatase (Boehringer-Manheim) and end labelled with g-^32P-ATP (ICN-Pharmaceuticals) 259 TBq/mmole, using T4 Polynucleotide Kinase (Boehringer Mannheim), according to the manufacturer's instructions. Band-Shift - The labelled fragments were incubated with male or female nuclear extracts for 15 min at room temperature followed by 15 min at 0 C, in a final voume of 20ul. The binding buffer used contained 10mM HEPES pH 7.9, 25mM CaCl2, 110mM MgCl2, 1æg acetylated bovine albumin and 1mg of poly d(I)d(C) (Pharmacia). b-estradiol (SIGMA) and Tamoxifen (ICI), an estrogen antagonist, were incubated in the concentrations indicated in the Figures. After incubation, the samples were fractionated in a 4% non-denaturating polyacrylamide gel using 1X TAE buffer (0.04 M Tris-acetate, pH 8.0, 1mM EDTA). The gel was dried under vacuum and exposed to X-ray film with an intensifying screen. RESULTS Figure 1 shows the results obtained by incubating schistosome total protein extracts with ^3H -17-b-estradiol. A mean value of 5.8 fmol of estradiol/mg protein, was obtained using female protein extracts. Male protein extracts yielded a value of 4.1 fmol/mg protein. The differences between the two experimental groups were not significant, however. Ecdysones, which are known to occur in schistosomes (Nirde et al. 1983), were also tested in the steroid binding assay. The results are shown in Fig.2. Surprisingly, in the presence of 20-hydroxyecdysone, ^3H 17-b-estradiol binding to the receptors was potentiated several fold in both, male and female protein extracts. The increased binding of estradiol to the protein receptor may have resulted from an allosteric effect. Figure 3 shows the results of experiments measuring the effect of Tamoxifen and 17-b-estradiol on the mobility shift of the 5' 107 bp F-10 fragment, caused by male and female nuclear proteins. Female nuclear proteins bound to the 107 bp fragment with a more pronounced effect on the electrophoretic mobility of the complex than that observed for the male nuclear proteins (compare lane 2 with lane 7). Furthermore, nuclear proteins extracted from female worms which had been incubated with Tamoxifen failed to produce the retardation observed in lane 2. Although band shift of the 107 bp fragment did Ccur with the nuclear proteins of the Tamoxifen treated female worms, this became similar to that produced by nuclear proteins of male worms (compare lane 6 to lane 7). Tamoxifen treatment did not affect the mobility shift of the complex formed between male nuclear proteins and the 107 bp DNA fragment (compare lane 7 to lane 11), suggesting that the effect of Tamoxifen was directed specifically to female nuclear proteins. The results in Fig.3 also show that incubation of both live male and female worms with estradiol did not significantly affect the mobility shift caused by the nuclear proteins of either sex . The same type of experiment was carried out with the 3' end, 306 bp F-10 fragment, containing the steroid regulatory element. The results are shown in Fig.4. Again it can be seen that nuclear proteins obtained from female proteins produced a distinct pattern of binding, when compared to the male proteins (lane 2 and lane 7). Also, pre-treatment with Tamoxifen abolished the female-specific shift as shown in Fig.4, lane 6. Although the complex observed in lane 6 did not form a sharp band, it did not comigrate exactly with the complex produced by proteins from untreated male worms (lane 7), as observed for the 107 bp fragment. By comparing lane 7 to lanes 8, 9 and 10, it can also be seen that estradiol and Tamoxifen, albeit discretely, affected the mobility shift produced by the male protein extract. The effect of Tamoxifen was also studied in vivo, by injecting the drug into S.mansoni infected hamsters, extracting the nuclear proteins from adult worms and incubating them with the 107bp and 306 bp F-10 fragments for the band-shift experiments. The results are shown in Fig.5. It can be seen that Tamoxifen affected the interaction of proteins with the DNA in both preparations (lanes 3, 5, 8 and 10). Curiously, in these experiments, the gel shift retardation effect of both protein preparations (male and female), was abolished by Tamoxifen. However, using another approach, Tamoxifen was shown to exert its effect primarily on females. Two main features could be observed: (a) S. mansoni infected and - Tamoxifen injected hamsters, displayed a higher rate of immature females, as shown in Table; (b) The eggs of worms recovered from hamsters injected with the anti-estrogen presented gross morphological anomalies, as shown by the arrow in Fig.6. No noticeable effects of Tamoxifen on male parasites could be detected. In order to investigate whether the binding of the schistosome nuclear proteins to the F-10 fragments was dependent on the zinc finger structural motif of steroid receptors (containing thiol groups), the nuclear extract was chromatographed on a AFFI-GEL 501 affinity column, prior to the gel mobility assay. This resin displays affinity for thiol groups. The results obtained with the 5', 107 bp fragment are shown in Fig.7. It is clear that only those proteins retained by the AFFI-GEL column were able to bind to the DNA fragment (lanes 4 and 5). A similar result was obtained with with the 3' end, 306 bp fragment (Fig.8), where it is evident that only those proteins retained by the matrix produced a band shift (lane 5). When male and female nuclear proteins were chromatographed separately on the AFFI-GEL column, band shifts were observed mainly with retained proteins extracted from male worms, as shown in Fig. 9, lanes 10 and 11. In lane 9, the 107 bp fragment was shifted to the same position after incubation with the unbound proteins. The absence of a shift observed with both female, bound and unbound proteins (Fig. 9, lanes 3- 6) was interpreted as due to loss of protein during the chromatographic procedure. Similar results were obtained with the 3'end fragment, as shown in Fig.10. DISCUSSION The ligand binding experiments using crude extracts of male and female schistosomes incubated with ^3H 17-b-estradiol and 20-hydroxyecdysteroid suggested the occurrence of steroid receptors. The apparent potentiation of 20-hydroxyecdysone on estradiol binding indicated that the schistosome putative estrogen receptor could be modulated by other steroids. Using the gel mobility assay we were able to establish that proteins in nuclear extracts of adult schistosomes, bound to the gene F-10. Among these proteins, some were only found in female nuclear protein extracts. The binding of female schistosome nuclear proteins to the fragments corresponding to the 5' and 3' ends of the gene F-10 was shown to be influenced by Tamoxifen, an estrogen antagonist both, in vitro and in vivo. In vitro, this effect of Tamoxifen was restricted to the female protein extracts. On the other hand, Tamoxifen injected into hamsters was shown to affect both, the development of females and of eggs, as indicated by pronounced morphological alterations. These effects of Tamoxifen could have been explained by inhibition of protein biosynthesis, a known property of the anti-estrogen. However, control experiments (not shown) failed to demonstrate a decreased incorporation of 35S-methionine onto proteins of Tamoxifen-treated parasites, thus suggesting that the drug may have exerted its effect by blocking the access of the schistosome nuclear proteins to steroid receptors. Other results strenghtening the argument for the occurrence of steroid receptors among the nuclear proteins, were obtained from the affinity chromatography experiments using the AFFI-GEL matrix. These experiments showed that the binding of the proteins to the different fragments of the F-10 gene, seem to require thiol groups, and therefore, clusters of cysteine residues. The occurrence of these cysteine clusters is consistent with the occurrence of zinc-fingers, a structural feature of steroid receptors. Whereas unpurified female and male purified cysteine containing proteins were clearly able to produce a shift with the 5' and 3' fragments, this effect was not observed with either the retained and unretained fractions of the female protein preparation. Taken together, the results in this paper make a case for the occurrence of steroid receptors in schistosomes.
TABLE Effect of Tamoxifen on the maturation of female Schislosoma mansoni ----------------------------------------------- % of immature females Tamoxifen 54 Control 8Schistosoma mansoni infected hamsters were injected daily with 100ug of Tamoxifen begining at five weeks after infection. The worms were perfused and observed in a light microscope at low magnification. The number of stunted worms lacking vitelline glands was then counted. ACKNOWLEDGEMENTS To expert technical assistance of Mrs Sonia Cristina Freire and Ms Maria Marta Freire. REFERENCES Bobek LA, Lo Verde PT, Rekosh DM 1989. Schistosoma haematobium: Analysis of eggshell protein genes and their expression. Exp Parasitol 68: 17-30. Bobek LA, Rekosh DM, Lo Verde PT 1991. Schistosoma japonicum: Analysis of eggshell protein genes, their expression and comparison with similar genes from other schistosomes. Exp Parasitol 72: 381-390. Chen L, Rekosh DM, Lo Verde PT 1992. Schistosoma mansoni p48 eggshell protein gene: characterization, developmentally regulated expression and comparison to the p14 eggshell protein gene. Mol. Bi Chem Parasitol 52: 39-52. Engelender S, Giannini ALM, Rumjanek FD 1993. Protein interactions with a gender-specific gene of Schistosoma mansoni: characterization by DNase I footprinting, band- shift and UV cross-linking. Mol Cell Bi Chem 124: 159-168. Henkle KJ, Cook GA, Foster LA, Engman DM, Bobek LA, Cain GD, Donelson JE 1990. The gene family encoding eggshell proteins of Schistosoma mansoni. Mol Bi Chem Parasitol 42: 69-82. Kunz W, Opatz K, Finken M, Symmons P 1987. Sequence of two genomic fragments containing an identical coding region for a putative eggshell precursor protein of Schistosoma mansoni. Nucl Acids Res 15: 5894. Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ 1951. Protein measurement with the Folin phenol reagent. J Biol Chem 193: 265-275. Nirde P, Torpier G, De Reggi ML, Capron A 1983. Ecdysone and 20-hydroxyecdysone: New hormones for the human parasite Schistosoma mansoni. FEBS Letters 151: 223-227. Rickwood D, Hames BD 1990. Gel electrophoresis of nucleic acids, a practical approach, p. 225-248. In GH Goodwin, The analysis of sequence-specific DNA-binding protein in cell extracts, Oxford University Press, New York. Rumjanek FD, Braga VMM, Kelly C 1989. DNA binding proteins of Schistosoma mansoni recognizing an hexanucleotide motif Ccuring in genes regulated by steroids. Comp Bi Chem Physiol 94: 807-812. Simpson AJG, Chaudhri M, Knight M, Kelly C, Rumjanek FD, Martin S, Smithers SR 1987. Characterization of the structure and expression of the gene encoding a major female specific polypeptide of Schistosoma mansoni. Mol Biochem Parasitol 22: 169-176. Smithers SR, Terry RJ 1965. The infection of laboratory hosts with cercariae of Schistosoma mansoni and the recovery of adult worms. Parasitology 55: 695-700. Copyright 1995 Fundacao Oswaldo Cruz (FIOCRUZ) The following images related to this document are available:Halftone images[oc95037h.gif] [oc95037f.gif] [oc95037e.gif] [oc95037i.gif] [oc95037j.gif] [oc95037c.gif] [oc95037d.gif] [oc95037g.gif]Photo images[oc95037g.jpg] [oc95037i.jpg] [oc95037c.jpg] [oc95037j.jpg] [oc95037d.jpg] [oc95037h.jpg] [oc95037f.jpg] [oc95037e.jpg]Line drawing images[oc95037b.gif] [oc95037a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}