 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias Instituto Oswaldo Cruz, Vol. 90(2):195-204 mar./apr. 1995 The Snail Hosts of Schistosomiasis: Some Evolutionary and Ecological Perspectives in Relation to Control JD Thomas School of Biological Sciences, University of Sussex, Falmer, Brighton, BN1 9QG, UK
Code Number: OC95040
Size of Files:
Text: 48K
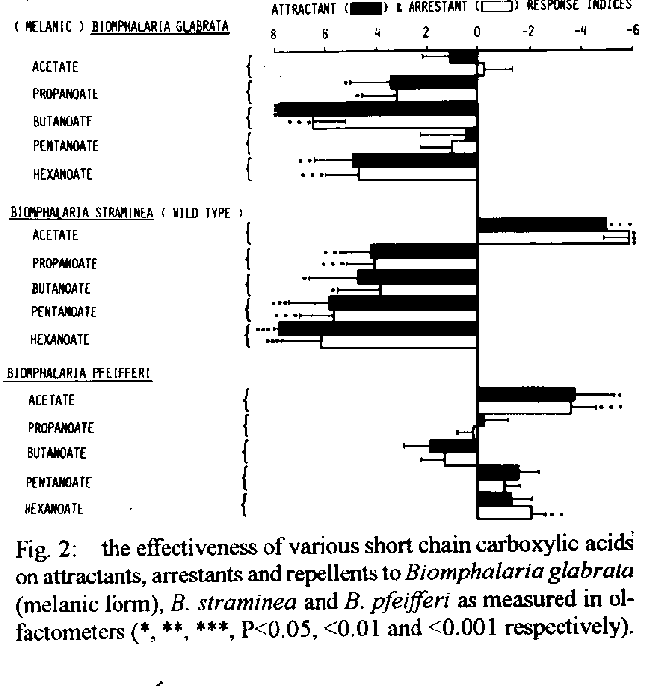
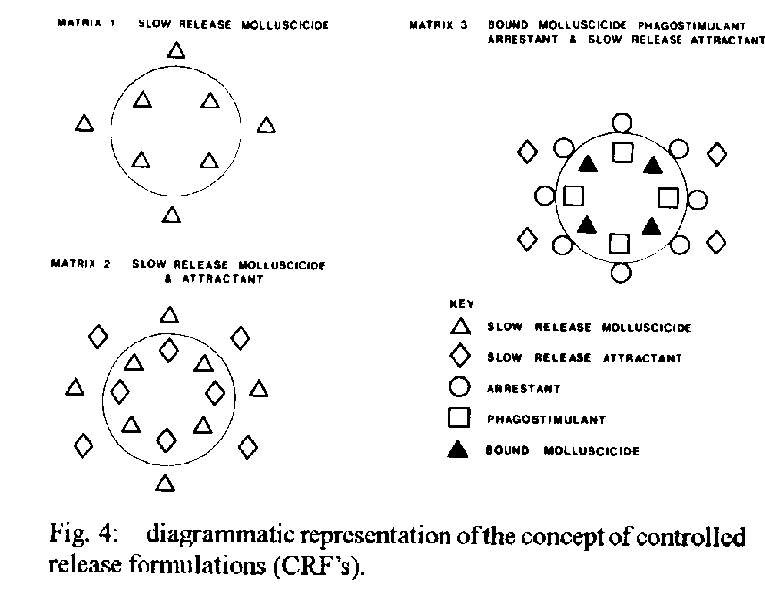
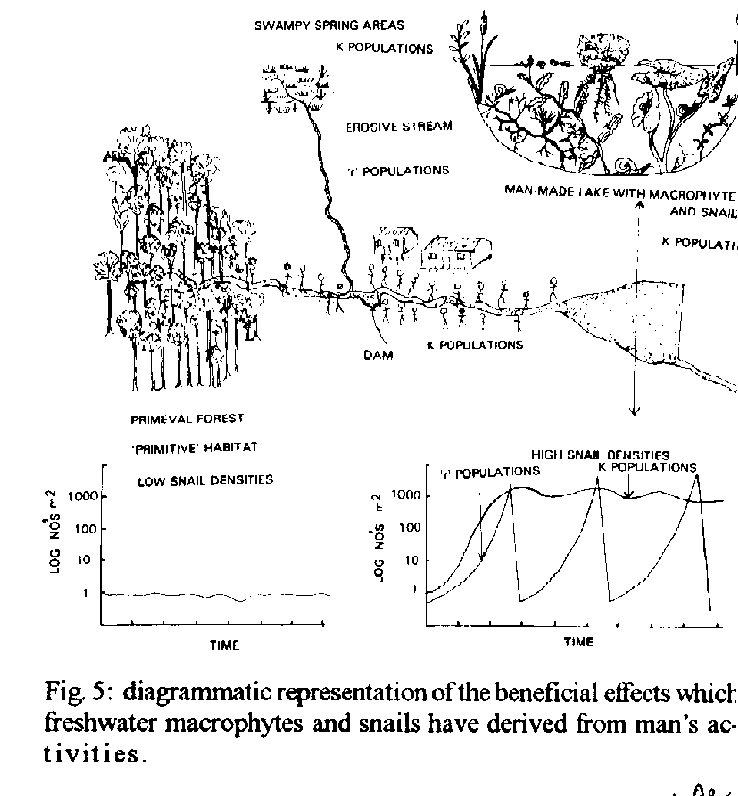
Graphics: Line Drawings (gif) - 74KDespite opportunities for radiation provided by spatio- temporal isolation, the basic morphological plan of pulmonate snails has remained conservative. In consequence of the resulting dearth of morphological characters and their plasticity, there is a case for using biochemical characters such as exogenous chemicals released by the snails (e.g. amino acids) and their chemoreception niche as taxonomic aids to classify snails of medical importance. As these same chemicals are used by snails to distinguish conspecifics they could also be used as environmental antibodies in controlled release formulations (CRF s) designed to remove target snails in a specific, cost-effective and ecologically acceptable manner. The snails, surface-living bacteria, algae and macrophytic plants are considered as co-evolved, interactive modular systems with strong mutualistic elements. Recently, anthropogenic perturbations such as deforestation, and damming of flowing waters, have benefited these modules whereas others such as river canalization, acid deposition, accumulation of pesticide residues and eutrophication have harmed them. Research is needed to elucidate the factors which limit the growth of snails in primitive habitats, uninfluenced by man, as well as in those subject to harmful anthropogenic factors. The understanding thus gained could be applied to develop cost-effective primary health care strategies to reduce or prevent transmission of schistosomiasis and other water related diseases. Key words: pulmonate-molluscs - evolution - taxonomy - controlled release formulations - control The gastropod molluscs, which first appeared in the fossil records in the late Cambrian or Ordovician periods (500 - 600 million years ago), were sediment dwelling organisms with radulae which were beautifully adapted for raking the highly productive epilithon or aufwuchs on the sediment surface of the shallow, eutrophic, near-shore waters (McMahon et al. 1974, Russell-Hunter 1978). Representatives of the super-order Pulmonata, to which the snail hosts of schistosomiasis in South America and Africa belong, first appeared in the Devonian period (Morton 1955). According to Ellis (1926), Morton (1955), McMahon (1983), the evolutionary history of the pulmonate line was characterized by continual invasion of successively higher and more aerial levels of estuaries and intertidal shores, a mode of life which eventually led to a totally terrestrial air-breathing and reproductive schesis. The freshwater pulmonates which evolved from these terrestrial ancestors and began to invade freshwaters in the Jurassic period (180 million years ago) were even then represented by families and genera which are assignable to those occupied by present day freshwater pulmonates (McMahon 1983). It appears, therefore, that the pulmonate snails have been remarkably conservative with regard to their basic morphological plan despite the opportunities for radiation offered by spatio- temporal isolation provided by the discrete nature of water bodies and the separation of land masses resulting from continental drift. Thus, despite the fact that the land masses which constitute South America and Africa to-day began to move apart during the Jurassic period, many of the extant pulmonate snail faunas of the two continents share the same families and generic names. One interesting exception is the sub-family Bulininae which is restricted to Africa. In consequence of their medical and veterinary importance as hosts of trematode parasites, the taxonomy of pulmonate snails has been thoroughly investigated, both in Africa (Mandahl Barth 1958, Brown 1980) and in South America (Barbosa et al. 1968) using classical taxonomic methods, based on visual, morphological characters. Unfortunately, some of the morphological characters, particularly those related to the shell, have limited value as their expression can be influenced by environmental factors (Brown 1980, Madsen 1984, Pieri & Thomas 1992). There is, therefore, a general consensus among molluscan taxonomists that there is a need to re-evaluate existing taxonomies using as wide a range of characters as possible, including those of a cytological and biochemical nature. To date, attention has been focussed mainly on specific gene products such as endogenous isoenzymes using electrophoretic and immunological methods (Brown 1980). Ultimately, of course, it is hoped that the use of DNA sequencing techniques will provide the definitive answers although many technical difficulties remain to be overcome. Unfortunately, the possibility of using exogenous biochemical factors of snail origin as taxonomic characters seems to have been largely ignored. This is surprising when it is considered that pulmonate snails use the chemical sense as the major modality and identify their conspecifics, for mating purposes, on the basis of chemical information provided by specific chemicals, or medleys of chemicals, of snail origin which act as pheromones (Thomas 1982). It follows, therefore, that exogenous chemicals of snail origin should have a great deal of information to the taxonomist as well as to snails. Some support for this hypothesis is provided by observations that pulmonate snails follow mucus trails deposited by their conspecifics to a significantly greater extent than those deposited by other species (Bousfield et al. 1981). Wright (1964) has also shown that different genetic races of Lymnaea peregra could be distinguished on the basis of unidentified fluorescent substances in their mucus. Subsequent technological developments, such as High Performance Liquid Chromatography (HPLC), have made it possible to identify specific exogenous organic chemicals released by the snails. By using HPLC and cluster analysis, it has been shown recently that various species of African and American pulmonates, including those that serve as hosts of schistosomiasis, can be characterized on the basis of the amino-acid medleys released by them under standard conditions (Fig. 1, Thomas & Eaton 1993). This result is not surprising as it is to be expected that their protein metabolism, which determines the amino-acid release pattern of particular snail species, will have some unique, genetically determined features. In this connection it is of interest that cluster analysis of amino-acid patterns reveals a high degree of similarity between the South American genotypes investigated (albino B. glabrata, melanic B. glabrata and B. tenagophila) whereas, in contrast, the South American and African species investigated were very dissimilar. This divergence between the South American and African species in their amino-acid medleys mirrors that found when their short-chain carboxylic acid chemoreception niches were compared. Thus the short-chain carboxylic and chemoreception niches of B. glabrata and B. straminea, another South American pulmonate snail, are very similar whereas both of these species have short- chain carboxylic and chemoreception niches which are very different from that of B. pfeifferi, an African species (Thomas et al. 1983, Fig. 2). These biochemical differences between the African and South American pulmonates are not surprising when it is considered that they have been geographically separated since the Jurassic or Cretaceous period. The hypothesis that snail genotypes might be distinguished on the basis of their exogenous biochemical milieu and chemoreceptors is worthy of further investigation, not only for taxonomic reasons but also because the information which would be gained from such research has application in the design of controlled release formulations (CRF's). This concept, which has been described by Thomas et al. (1980), involves using ingestible microcapsules (62 - 125uM in diameter) capable of releasing species-specific attractants, arrestants and phagostimulants externally in the snail s environment and a toxicant in the gut of the snail following ingestion (Fig. 3). Attractants, such as propanoate or butanoate, and phagostimulants, such as maltose, have proved very efficacious when incorporated into protein microcapsules and tested under laboratory conditions. Such microcapsules, containing a natural product such as saponin from the Soap- Berry plant (Phytolacca dodecandra) or the Alligator weed (Alternanthera sessilis) (Lema et al. 1984, Thomas 1987) could conceivably be used to remove target snails in an ecologically acceptable and cost-effective manner. With the technology currently available these microcapsular CRF s, or environmental antibodies as they might be described, could be developed at relatively little cost. In contrast, research aimed at producing vaccines against the adult schistosomes has proved to be very costly and unproductive due to the serious problems associated with the development of vaccines. These stem from the fact that many people, particularly children, are highly susceptible to re-infection after chemotherapeutic cure (Dunne et al. 1987). As there appears to be no long-term acquired immunity to the parasites, at least in children, immunologists will have to improve on what nature has been unable to accomplish by natural selection. This is clearly a difficult task as the parasites, which have evolved in the host s immune system, have developed many sophisticated strategies to inactivate and even make use of the host s immune mechanisms to enhance their survival (Dunne et al. 1987). PULMONATE SNAILS: CO-EVOLUTION AND BIOENGINEERING APPROACHES TO SNAIL CONTROL Freshwater pulmonate snails, which evolved from terrestrial ancestors, are represented to-day by a continuum commencing with amphibious species living in the ecotone between land and water (e.g. Lymnaea truncatula), through intermediately adapted forms such as the Physidae to those which are more committed to life in water (e.g. some species of the Planorbidae). Although amphibious snails such as L. truncatula are often strongly associated with particular species of emergent macrophytes, which act as bioindicators of their presence (Moens 1981) there are no direct linkages. Instead the snails interact with epilithic algae and bacteria living on the surface of the sediments. This system, consisting of three interactive components and three sub-sets, has been described as Module Type 1 (Thomas 1990) (Fig. 4). In contrast, pulmonate snails, which have become more committed to life in freshwaters, interact to a much greater extent with species of macrophytes which have also become fully aquatic, as well as their epiphytic algae and bacteria. Such a system, therefore, has four components and six sub-sets and has been described as Module Type 2 (Thomas 1990) (Fig. 4). It is likely that the associations described in Module Type 2 first developed in the late Jurassic or Cretaceous period when both macrophytes (Scagel et al. 1965) and pulmonate snails (Russell-Hunter 1978) were beginning to invade freshwaters from the land. As was the case with the pulmonate snails certain of the aquatic angiosperm taxa became highly adapted morphologically for their new habitat by the Cretaceous and many modern families and even genera were recognizable in the fossil records for this period (Scagel et al. 1965). Analysis of all the sub-sets in Modules Type 1 and 2 reveal evidence of mutualistic elements which result in benefits to the interacting components. The Module Type 2 concept has been tested by reference to the following hypothesis and propositions: The associations between components of the module including pulmonate snails and macrophytes should be strong and persistent - In seasonally drying habitats it has been shown that the pulmonate snail hosts of schistosomiasis are often significantly associated with emergent, amphibious plants such as species of Commelina, Paspalum, Acroceras, Alternanthera, Cyperus, Glyceria and Juncus (Ndifon 1979, Moens 1981, Pieri 1985, Pieri & Thomas 1987, Ndifon & Ukoli 1989). In such habitats the snails tend to function as components of Module Type 1 and may even emigrate from water in response to a fall in temperature which acts as a predictive signal for the dry season (Pieri & Thomas 1992). In more permanent water bodies, including those resulting from man's activities (e.g. irrigation systems, man-made lakes) the snails such as Biomphalaria or Bulinus are commonly found living on the surfaces of specialized aquatic plants with submersed or floating leaves, such as Potomogeton crispus, Eichornia crassipes, Nymphaea lotus, Ceratophyllum demersum and Lemna paucicostata (Dazo et al. 1966, Thomas & Tait 1984, Paperna 1969, Odei 1973, Ndifon 1979, Lodge 1985, 1986). The- se linkages tend to be stronger when both plants and snails are highly committed to a fully aquatic existence (e.g. Ceratophyllum demersum and Bulinus forskali) (Paperna 1969, Odei 1973). The more vulnerable elements in the modular system will have evolved sophisticated mechanisms to minimize exploitation by potential grazers - This hypothesis is supported by observations that aquatic pulmonate snails feed almost entirely on senescent macrophyte tissue (detritus) and epiphytic algae rather than on living macrophyte tissue (Boycott 1936, Reavell 1980, Thomas et al. 1985, Underwood 1989). There is also evidence that some species of epiphytic algae have evolved strategies to resist ingestion and digestion by snails and, as a result, may benefit from the presence of snails (Thomas 1990, Underwood & Thomas 1990). Growth, reproduction and longevity of interactive components in the module should be enhanced by co-existence - This hypothesis also receives support from empirical evidence. Thus, the growth, sexual reproduction and survival of the specialized submersed aquatic macrophyte, Ceratophyllum demersum, are significantly enhanced by the presence of pulmonate snails (BrÆnmark 1985, 1989, Underwood 1989, 1991, Thomas 1990). The converse proposition, namely that snails benefit from the presence of plants, is also supported by experimental evidence (Pimentel & White 1959a, b, Van Schayck 1985, 1986, Daldorph & Thomas in press). These observations raise another important question: as the pulmonate snails ingest very little living macrophyte tissue and most of the algae and bacteria which are ingested also remain viable, how do the snails satisfy their nutritional requirements? Evidence presented by Thomas (1990) indicate that the snails derive some of their energy by taking up photoassimilates released by living plants. However, it seems likely that short chain carboxylic acids (C2 - C5) may constitute one of the major components of their diet. These are released by anoxic cores within decaying organisms (Patience et al. 1983, Sterry et al. 1985) and may be accumulated either transintegumentally or orally, together with detritus, by the snails (Thomas et al. 1984, 1990, Thomas 1990). As these acids, and also sugars, released as photoassimilates by plants serve as attractants they can be incorporated into CRF's designed to remove target snails selectively (Fig.3). THE INFLUENCE OF MAN ON SNAIL HABITATS Perturbations which have been favourable to the snails - Before the exponential increases in the population densities of modern man (Homo sapiens sapiens) began it is likely that the availability of habitats for aquatic macrophytes, surface living algae and pulmonate snails would have been greatly reduced by light attenuation caused by high riparian forests (Fig. 5). Such primitive habitats still exist in the National Parks in Minas-Gerais, Brazil (Freitas 1976). As the pulmonate snails have evolved in these primitive habitats for more than a hundred million years it would be expected, on the basis of conventional ecological knowledge, that their numbers would have been regulated at low levels by pathogens and predators. According to Pimentel and White (1959b) and Freitas (1976) the latter include small rodents which are well represented both in terms of species and numbers in high forests. The expansion in human population densities has been accompanied by many anthropogenic manipulations which have benefitted aquatic macrophytes and pulmonate snails. In the developing countries these included deforestation, the formation of man-made lakes and ponds, small scale damming of streams and the provision of water supplies without accompanying drainage in urban areas (Fig. 5). Typically lotic habitats in these areas can be divided into three distinct zones: (i) Springs and marshy areas in the headwater zone where the snail populations exhibit stable or 'K' characteristics (Pieri 1985); (ii) More erosive lotic habitats where the gradients are steeper. Here the snail populations show 'r' characteristics, the densities being high in deeper, isolated pools in the dry season and very low during the wet season (Fig. 5); (iii) More low-lying areas, with less steep gradients, where the streams have substantial amounts of depositing sediments and are often dammed by the construction of small weirs. In such habitats the snail populations may often exhibit 'K' characteristics. The construction of small ponds or lakes generally results in conditions which are ideal for aquatic macrophytes and molluscs, particularly if they are subject to low or moderate levels of nutrient loading. Further research to identify the factors which disadvantage the snails in primitive habitats dominated by high forests and also in erosive stream habitats is clearly needed as the results could have application in the development of snail control strategies. The effects of shading by trees could be investigated experimentally by using opaque polythene sheeting. Recent studies using such material have shown that the resulting changes are indeed detrimental to many pulmonate snail species (Thomas & Daldorph 1991). The classical approach of identifying the predators or pathogens which serve as biocontrol agents in the primitive or original habitats, and then introducing them in new habitats where a prey species has reached epidemic or pest proportions has been used successfully on many occasions by agricultural entomologists. A similar approach by malacologists might well prove to be cost-effective. The significant negative relationships between snail densities and both water velocity and slope is well documented (Harry & Cumbie 1956, Pimentel & White 1959a, Scorza et al. 1961, Etges & Frick 1966) although the causative mechanisms remain to be elucidated. Dislodgement by fast currents is clearly a factor and Jobin and Ippen (1964) have shown that B. glabrata was dislodged from a sand-silt substrate when the current exceeded 30cm sec^-1. It is also tacitly assumed that the clays or fine silt, brought into suspension by fast currents, are detrimental because of their mechanical effects. However, recent investigations have shown that certain kinds of colloidal clay may be harmful to snails as they adsorb heavy metals, such as copper, in a base rich external environment but then shed the metal as the toxic Cu^2+ ion when the clay particles enter the more acidic environment in close proximity to the snail's epithelium (Al-Sabri et al. 1993). Other possible harmful effects that may be caused by high flows include reduction in the availability of food items such as detritus, DOM and surface algae and the dilution of a growth promoting factor shown to be released by snails such as B. glabrata (Thomas et al. 1975a,b). As the snail populations in the more erosive stream habitats tend to be derived by seeding from 'K' populations in the headwater swamp areas, it is essential that measures directed at snail control should include the upstream refugia. ANTHROPOGENIC PERTURBATIONS LIKELY TO HAVE DETRIMENTAL EFFECTS ON FRESHWATER PULMONATE SNAILS Freshwater ecologists in the industrialized world are becoming increasingly concerned about the progressive decline in the biodiversity of freshwater communities. In some cases the changes have been very dramatic with weed-bed communities being replaced by phytoplankton (Moss 1983). Some of the mechanisms which may have been responsible for this decline in biodiversity in general, and the loss of the pulmonate snail fauna in particular, are discussed below. Canalization of rivers - This measure, which stems from pressure to reduce flooding in times of spate, is followed by a catastrophic decline in the area of depositing sediments and hence in the biomass and diversity of aquatic macrophytes and invertebrates, including pulmonate snails. In contrast, prosobranch molluscs, such as Potamopyrgus jenkinsi often flourish on eroding substrate in streams and may reach very high population densities. It is possible, therefore, that river canalization has been a major factor in facilitating the recent colonization of freshwaters by this hitherto estuarine species. These prosobranch molluscs are pre-adapted for existence on eroding substrates in flowing water by virtue of their relatively heavy, streamlined conical shell and parthenogenetic, viviparous life history strategy. Their mouth parts also allow them to exploit food items in the interstices between the gravel as well as to graze on epilithic algae on stone surfaces. As a result, prosobranchs appear to have a competitive advantage over pulmonate snails when the substrate is predominantly erosive in character. It is not surprising, therefore, that there are many reports of prosobranch molluscs apparently displacing pulmonate snails in freshwater bodies as far afield as Europe (Leger & Leger 1974) and Brown (1980) and St Lucia (Jordan 1985). However, the question of whether the disappearance of pulmonate snails is due directly to interspecies competition by the prosobranchs or to other concomitant detrimental factors remains to be resolved. The latter may include the loss of macrophytes refugia with depositing sediments and high concentrations of ubiquitous pesticide residues. The question of whether pulmonate snails are more susceptible to the latter than prosobranch snails needs to be addressed. Acid deposition - The vulnerability of pulmonate molluscs, inhabiting water bodies with relatively low base cation concentrations, to anthropogenic acidification resulting from the burning of fossil fuel has been well documented by Okland (1990). This author observed that pulmonate snails disappear when the pH became less than 5.2. Although the actual mechanisms responsible for the demise of the snails remain to be elucidated, it is likely that the increase in the concentration of the toxic Al^3+ ion, resulting from the elevation in H^+ concentration, is the main proximate factor. However, a reduction in food availability for the snails may be an important secondary factor as it is known that the rate of decomposition of organic matter, and hence the rate of formation of short-chain carboxylic acids, will be impaired by the increased acidity (Chamier 1987, Palumbo et al. 1987). Increase in salinity - Endorheic salt lakes, formed by natural processes, were widespread before the advent of man on the evolutionary time scale (Moss 1988). However, the process of salination has been greatly accelerated in recent times by the expansion of irrigation practices which facilitate evaporative concentration of minerals. This process is of considerable economic concern because it has resulted in a significant decline in agricultural production. Estimates of the magnitude of the problem vary but according to Flowers et al. (1986) there are between 3.4 and 9.5 x 10^6 km^2 of salt affected land. To date attention has been focussed on the agricultural and economic consequences of salination. These can be extremely serious and according to Jacobsen and Adams (1958) the decline of the Sumeran civilization was attributable to this phenomenon. It is to be expected that the increased levels of salination will also cause a reduction in the biodiversity of the aquatic communities in the irrigated systems. From the public health viewpoint this could be beneficial as the pulmonate snail hosts of schis- tosomiasis are very intolerant to high salinities. Thus, according to Pimentel and White (1959a) B. glabrata in Puerto Rico does not occur in water bodies with a salinity greater than 5 - 10% sea water (~1000 - 2000 us). In the laboratory this species was found to be tolerant to water with a salinity of 20% sea water for a period of up to 35 days. These tolerance values are in accord with those for Bulinus bavayi (~1000 us) and Bulinus liratus (1500 - 2100 us) in Madagascar (Degremont 1973, cited in Brown 1980). In contrast other pulmonate snails, such as Bulinus obtusispira, have a much lower level of tolerance (< 600 us) whereas the prosobranch Melanoides tuberculata has a very wide tolerance range (200 - 20,000 us). It is to be expected, therefore, that Melanoides, which has an estuarine origin, would have a competitive advantage over the pulmonate snail hosts of schistosomiasis in water bodies devoid of vegetation and subject to wide-ranging salinities. Eutrophication and accumulation of pesticide residues - Eutrophication, due to increased inputs of nitrate and phosphates, is becoming an ever-increasing problem in both industrialized and developing countries. It has been demonstrated that this phenomenon can cause considerable changes in the community structure of many water bodies. In shallow, lentic water bodies eutrophication has been followed by a replacement of macrophytes by phytoplankton (Moss 1983, Daldorph & Thomas 1991) and a reduction in the numbers and diversity of fish and invertebrates in general (Mason & Bryant 1975, Moss 1983) and freshwater snails in the temperate zone in particular (Daldorph & Thomas 1991, Thomas & Daldorph 1994). There is also anecdotal evidence that tropical species of pulmonate snails such as B. truncatus, the snail host of S. haemotobium in Egypt and the Sudan may be adversely affected by increasing levels of eutrophication although B. alexandrina may be benefited (F Yussif & AA Daffola pers. comm.). Pesticide and herbicide loading of water bodies generally proceeds in parallel with nutrient loading in intensively farmed, irrigated areas. It is likely, therefore, that these will act synergistically with the nutrients to the detriment of the macrophyte beds and the snail fauna. The rationale for this hypothesis is that the herbicides may harm the macrophytes directly while the pesticides will harm not only the zooplankton, which control the phytoplankton, but also the snails which benefit the macrophytes by grazing the epiphytes. As a result, it would be predicted that a water subject to both nutrient and pesticide loading would quickly become phytoplankton-dominant and devoid of macrophytes and snails. Evidence in support of this hypothesis is provided by experiments recently carried out in the Lewes Brooks, UK (Daldorph & Thomas in press). In the temperate, industrialized countries, a great deal of research effort is directed at evaluating measures to reduce eutrophication and to restore aquatic habitats to their original state. However, as eutrophication may bring public health benefits in the developing countries it is arguable that control of water related diseases should take precedence over conservation issues. If this argument is accepted then so far as eutrophication is concerned there is a case for evaluating the efficacies of the following perturbations with a view to excluding the snail hosts and macrophytes in the sites of schistosomiasis transmission: (i) The long term effects of increased nutrient loading on the snail hosts of schistosomiasis, macrophytes, mosquito larvae and other members of the aquatic community under experimental conditions; (ii) Increased nutrient loading in combination with molluscicide application. As the relationship between snails and macrophytes is essentially mutualistic (Thomas 1990) it would be predicated that the simultaneous application of these two measures would accelerate the switch to stable algal-dominated systems; (iii) Increasing the density of zooplanktivorous fish to control the zooplankton. There are several other potential public health benefits from this approach. Firstly, phytoplankton in the water bodies could be exploited efficiently by suitable species of Tilapia thus providing people with a valuable source of protein in areas where protein dietary deficiency is commonplace. Secondly, the larvae of mosquitoes, which are involved in the transmission of a large number of water- related diseases, including malaria in the adult stage, are particularly vulnerable to predation in phytoplankton- dominated systems. CONCLUSIONS 1. The basic morphology of freshwater pulmonate snails has changed very little since they moved into freshwater from the terrestrial environment in the Jurassic despite opportunities offered by spatio-temporal isolation resulting from the discrete nature of water bodies and the separation of major land masses. Although the land masses, which constitute South America and Africa to-day, began to separate in the Jurassic their extant freshwater pulmonate snail faunas show close similarities at family and generic levels. 2. The dearth of reliable morphological characters and their environmental plasticity has obliged molluscan taxonomists to use endogenous biochemical factors, produced by specific genes such as isoenzymes, to resolve taxonomic problems. It is argued that this search for taxonomic characters should be extended to the active space surrounding pulmonate snails as this contains specific biochemical factors which are used by the snails to identify their conspecifics. Support for this suggestion is given by the observations that South American pulmonate snails can be discriminated from their congeneric African species on the basis of their amino acid medleys and short chain carboxylic acid chemoreception niches. 3. The identification of the biochemical factors in the active space of the snails could also be beneficial from the applied viewpoint. Thus, factors that serve as attractants and arrestants could be incorporated into environmental 'antibodies' in the form of controlled release microcapsules designed to remove target snails in a species-specific, ecologically acceptable and cost-effective manner. 4. Aquatic angiosperms and pulmonate snails have co-evolved as they both invaded freshwater habitats in the Jurassic. The snails, the macrophytes and their epiphytic algae and bacteria may constitute a four component, six sub-set modular system. Analyses of all the sub-sets reveal evidence of strong mutualistic interactions involving an exchange of metabolites between living components. The linkages within a module are strongest in the case of plants and snails which are fully committed to life in water (e.g. Ceratophyllum demersum and B. rohlfsi). Earlier anthropogenic influences, such as deforestation, increase in areas of depositing sediments by damming lotic systems, the construction of man- made lakes and irrigation systems and moderate eutrophication have benefitted the modules. This, in turn, has greatly facilitated the transmission of schistosomiasis. 5. In contrast, anthropogenic perturbations such as acid deposition, salination, intense eutrophication, accumulation of pesticide and herbicide resides, and canalization of rivers are all potentially harmful to pulmonate snails and other components of the module. Although from the conservation viewpoints such effects are rightly considered disastrous, there are public health benefits to be derived in developing countries where schistosomiasis and other water-related diseases are prevalent. 6. Research aimed at the elucidation of the mechanisms responsible for the absence or dearth of pulmonate snails in aquatic habitats within high forests (primitive habitats) as well as in those subject to harmful anthropogenic influences listed above is strongly indicated. The understanding thus gained could be used to develop primary health care management strategies aimed at preventing or reducing the transmission of schistosomiasis and other water-related diseases. It is arguable that the benefits to the communities in the developing countries of such an approach would far outweigh those that would be derived from the allocation of resources to the production of vaccines (if and when they are produced) and the continued use of drugs. ACKNOWLEDGMENTS To the Organizing Committee of the International Symposium on Schistosomiasis for the invitation to attend the symposium in Rio de Janeiro in December, 1993; to my friends at the Instituto Oswaldo Cruz, particularly Dr OS Pieri, for their hospitality; to the Royal Society, London, UK, for financial support; and to Mrs Pat Chatfield for typing the manuscript. REFERENCES Al Sabri IYN, Smith JD, Thomas JD 1993. Copper molluscicides for control of schistosomiasis: 3. Adsorption by clay suspensions. Env Sci & Technol 27: 299-303. Barbosa FS, Berry EG, Harry HW, Hubendick B, Malek EA & Paraense WL 1968. A guide for the identification of the snail intermediate hosts of schistosomiasis in the Americas. Scientific Publication No. 168. Pan American Health Organization, WHO, Washington, D.C. Bousfield JD, Tait AI, Thomas JD & Towner-Jones D 1981. Behavioural studies on the nature of stimuli responsible for triggering mucus trail following in the freshwater snail Biomphalaria glabrata (Say). Malacol Rev 14: 49-64. Boycott AE 1936. The habitats of freshwater molluscs in Britain. J Anim Ecol 5: 116 - 186. Bronmark C 1985. Interaction between macrophytes, epiphytes and herbivores: an experimental approach. Oikos 45: 26 - 30. Bronmark C 1989. Interactions between epiphytes, macrophytes and freshwater snails: a review. J Moll Stud 55: 299- 322. Brown DS 1980. Freshwater snails of Africa and their medical importance. Taylor & Francis, London. 487 pp. Chamier AC 1987. Effect of pH on microbial degradation of leaf litter in seven streams of the English Lake District. Oecologia 71: 491-500. Daldorph PWG & Thomas JD 1991. Evaluation of bioengineering approaches aimed at controlling pulmonate snails: The effects of light attenuation and mechanical removal of macrophytes. J Appl Ecol 28: 532-546. Daldorph PWG & Thomas JD (in press) Factors influencing the stability of freshwater macrophyte communities subject to nutrient enrichment: The role of sticklebacks, Pungitius pungitius, and freshwater snails. Freshw Biol. Dazo BC, Hairston NG & Dawood IK 1966. The ecology of Bulinus truncatus and Biomphalaria alexandrina and its implications for the control of Bilharziasis in the Egypt 49 Project Area. Bull WHO 35: 339-356. Degremont AA 1973. Mangoky Project; Campaign against schistosomiasis in the lower Mangoky (Madagascar), Basle, Swiss Tropical Institute. Dunne DW, Fulford AJC, Butterworth AE, Koech D & Ouma, H 1987. Human antibody response to Schistosoma mansoni: does antigen directed isotype restriction result in the production of blocking antibodies. Mem Inst Oswaldo Cruz, Rio de Janeiro 82, Suppl. IV: 101-104. Ellis RA 1926. British snails: a guide to the non-marine gastropods of Great Britain and Ireland, Pleistocene to Recent. Oxford University Press, London. 92 pp. Etges FJ & Frick LP 1966. An experimental field study of chemoreception and response in Australorbis glabratus (Say) under rheotactic conditions. Am J Trop Med Hyg 15: 434-438. Flowers TJ, Hajibagheri MA & Clipson NJW 1986. Halophytes. Quart Rev Biol 61: 313-337. Freitas JR de 1976. Ecologia de vetores de doen‡as, o habitat primitivo da Biomphalaria glabrata. Sept Rev Cienc Cult 28: 212-217.
Harry HW & Cumbie BG 1956. Stream gradient as a criterion of lotic habitats suitable for Australorbis glabratus in Puerto Rico. J Trop Med Hyg 5: 921-928. Jacobsen I & Adams RM 1958. Salt and silt in ancient Mesopotamian agriculture. Science 128: 1251-1258. Jobin WR & Ippen AT 1964. Ecological design of irrigation canals for snail control. Science 145: 1324-1326. Jordan P 1985. Schistosomiasis: The St Lucia project. CUP. Cambridge. 442 pp. Leger N & Leger P 1974. L extension de Potamopyrgus jenkinsi (Smith, 1989) en corse. Ann Parasit Hum Comp 49: 343-347. Lema A, Heyneman D & Silangwa SM 1984. Phytolacca dodecandra (Endod) Towards controlling transmission of schistosomiasis with the use of natural products. Final Report of the International Workshop, Lusaka, Zambia, March, 1983. Tycosly International Publishing Ltd., Dublin. 318 pp. Lodge DM 1985. Macrophyte-gastropod associations: observations and experiments on macrophyte choice by gastropods. Freshw Biol 15: 695-708. Lodge DM 1986. Selective grazing on periphyton: a determinant of freshwater gastropod microdistributions. Freshw Biol 16: 831-841. Madsen H 1984. Intraspecific and interspecific variation in shell morphology of Biomphalaria pfeifferi (Krauss, 1848) and Helisoma duryi (Wetherby, 1879) from Moshi, Tanzania, analysed by morphometric studies. J Moll Stud 50: 153-161. Mandahl-Barth G 1958. Intermediate hosts of Schistomosa: African Biomphalaria and Bulinus. WHO, Geneva. 89 pp. Mason CF & Bryant RJ 1975. Changes in the ecology of the Norfolk Broads. Freshw Biol 5: 257-270. McMahon RF 1983. Physiological ecology of freshwater pulmonates Vol. 6, Ecology p.359-461 In W D Russell- Hunter, The Mollusca, Academic Press, New York. McMahon RF, Hunter RD & Russell-Hunter WD 1974. Variation in aufwuchs at six freshwater habitats in terms of carbon biomass and of carbon:nitrogen ratio. Hydrobiologia 45: 391- 404. Moens J 1981. Les habitats de Lymnaea trunculata hote intermediaire de Fasciola hepatica. Rev Agric 34: 1563-1580. Morton JE 1955. The evolution of the Ellobiidae with a discussion on the origin of the pulmonata. Proc Zool Soc Lond 125: 127-168. Moss B 1983. The Norfok Broadlands: Experiments in the restoration of complex wetland. Biol Rev 58: 521- 561. Moss B 1988. Ecology of Freshwaters. Man and the Medium. Blackwells Sci Publ Oxford. Ndifon GT 1979. Studies on the feeding biology, anatomical variations and ecology of the vectors of Schistosomiasis and other freshwater snails in South Western Nigeria. PhD Thesis. University of Ibadan. 361 pp. Ndifon GT & Ukoli FMA 1989. Ecology of freshwater snails in South Western Nigeria. I. Distribution and habitat prefe- rences. Hydrobiologia 171: 231-253. Odei MA 1973. Observations on some weeds of malacological importance in the Volta Lake. Bull Inst fr Afr noire 35A: 57-66. Okland J 1990. Lakes and snails. Universal Book Services, Dr W Backhuys, Oegstgeest. 516 pp. Palumbo AV, Mulholland PJ & Elwood JW 1987. Microbial communities on leaf material protected from macroinvertebrate grazing in acidic and circumneutral streams. Can J Aquat Sci 44: 1064-1070. Paperna I 1969. Aquatic weeds, snails and transmission of Bilharzia in the new man-made Volta Lake in Ghana. Bull Inst fr Afr noire 31A: 487-499. Patience RL, Sterry PR & Thomas JD 1983. Changes in the chemical composition of a decomposing aquatic macrophyte Lemna paucicostata. J Chem Ecol 9: 889- 991. Pieri OS 1985 Studies on the host snails of schistoso- miasis from North East Brazil, with special reference to diapause in Biomphalaria glabrata (Say). PhD Thesis, University of Sussex. 155 pp. Pieri OS & Thomas JD 1987. Snail-host control in the eastern coastal areas of North East Brazil. Mem Inst Oswaldo Cruz 82: 197-201. Pieri OS & Thomas JD 1992. Induction of morphological, behavioural and physiological changes in polymorphic populations of Biomphalaria glabrata (Say) by an environmental factor of predictive value. J Med & Appl Malac 4: 43-56. Pimentel D & White PC 1959a. Physicochemical environments of Australorbis glabratus, the snail intermediate host of Schistosoma mansoni in Puerto Rico. Ecology 40: 533-541. Pimentel D & White PC 1959b. Biological environments and habits of Australorbis glabratus. Ecology 40: 541 - 550. Reavell PE 1980. A study of the diet of some British freshwater gastropods. J Conchol 30: 253 - 271. Russell-Hunter WD 1978. Ecology of freshwater pulmonates, p.344-383. In V Fretter & J Peake, Pulmonates, Academic Press, London. Scagel RF, Bandoni JR, Rouse GE, Schofield WB, Stein JR & Taylor, TMC 1965. An evolutionary survey of the plant kingdom. Blackie, London. Scorza JV, Silva J, Gonzales L & Machado R 1961. Stream velocity as a gradient in Australorbis glabratus (Say, 1818). Z Tropenmed Parasit 12: 191-196. Sterry PR, Thomas JD & Patience RL 1985. Changes in the concentrations of short-chain carboxylic acids and gases during decomposition of the aquatic macrophytes Lemna paucicostata and Ceratophyllum demersum. Freshw Biol 15: 139-153. Thomas JD 1982. Chemical ecology of the snail hosts of Schistosomiasis: snail-snail and snail-plant interactions. Proc 7th Europ Malac Congr Malacologia 22: 81- 91. Thomas JD 1987. An evaluation of the interactions between freshwater pulmonate snail hosts of human schistosomes and macrophytes. Phil Trans R Soc Lond B315: 75-125. Thomas JD 1990. Mutualistic interactions in freshwater modular systems with molluscan components. Adv Ecol Res 20: 125-178. Thomas JD, Cowley C & Ofosu-Barko J. 1980. Behavioural responses to amino and carboxylic acids by Biomphalaria glabrata, one of the snail hosts of Schistosoma mansoni, p.433-448. In R Baker, Controlled release of bioactive materials. Academic Press, New York. Thomas JD & Daldorph PWG 1991. Evaluation of bioengineering approaches aimed at controlling pulmonate snails: The effects of light attenuation and mechanical removal of macrophytes. J Appl Ecol 28: 532 - 546. Thomas JD & Daldorph PWG 1994. The influence of nutrient and organic enrichment on a community dominated by macrophyte and gastropod molluscs in a eutrophic drainage channel: Relevance to snail control and conservation. J Appl Ecol 31: 571-588. Thomas JD & Eaton P 1993. Amino acid medleys of snail origin as possible sources of information for conspecifics, Schistosome miracidia and predators. Comp Biochem Physiol 106C: 781-796. Thomas JD & Tait AI 1984. Control of the snail hosts of Schistosomiasis by environmental manipulation: A field and laboratory appraisal in the Ibadan area of Nigeria. Phil Trans R Soc Lond B305: 201-253. Thomas JD, Goldsworthy GJ & Aram RH 1975a. Studies on the chemical ecology of snails. The effect of chemical conditioning by adult snails on the growth of juvenile snails of the same species. J Anim Ecol 44: 1-27. Thomas JD, Kowalczyk C & Somasundaram B 1990. The chemical ecology of Biomphalaria glabrata, a freshwater pulmonate mollusc: the uptake and assimilation of exogenous glucose and maltose. Comp Biochem Physiol 95A: 511- 528. Thomas JD, Lough AD & Lodge RW 1975b. The chemical ecology of Biomphalaria glabrata (Say) the snail host of Schistosoma mansoni (Sambon): the search for factors in the media conditioned by snails which inhibit their growth and reproduction. J Appl Ecol 12: 421-437. Thomas JD, Nwanko DI & Sterry PR 1985. The feeding strategies of juvenile and adult Biomphalaria glabrata (Say) under simulated natural conditions, and their ecological significance. Phil Trans R Soc B226: 177-209. Thomas JD, Ofosu-Barko J & Patience RL 1983. Behavioural responses to carboxylic and amino acids by Biomphalaria glabrata (Say), the snail host of Schistosoma mansoni (Sambon) and other freshwater molluscs. Comp Biochem Physiol 75C: 57-76. Thomas JD, Patience RL & Sterry PR 1984. Uptake and assimilation of short chain carboxylic acids by Biomphalaria glabrata (Say) the freshwater pulmonate snail host of Schistosoma mansoni (Sambon). Proc R Soc Lond B222: 447-476. Underwood GJC 1989. Interactions between freshwater pulmonate snails, macrophytes and epiphytes. PhD Thesis, University of Sussex. 218 pp. Underwood GJC 1991. Growth enhancement of the macrophyte Ceratophyllum demersum in the presence of snail Planorbis planorbis: The effects of grazing and chemical conditioning. Freshw Biol 26: 325 - 334. Underwood GJC & Thomas JD 1990. Grazing interactions between pulmonate snails and epiphytic algae and bacteria. Freshw Biology 23: 505 - 522. Van Schayck CP 1985. Laboratory studies on the relation between aquatic vegetation and the presence of two Bilharzia- bearing snail species. J Aquat Pl Manag 23: 87- 91. Van Schayck CP 1986. The effect of several methods of aquatic plant control on two Bilharzia-bearing snail species. Aquat Bot 24: 303-309. Wright CA 1964. Biochemical variation in Lymnaea peregra (Mollusca: Basommatophora). Proc Zool Soc Lond 142: 371-378. Copyright 1995 Fundacao Oswaldo Cruz (Fiocruz)
The following images related to this document are available:Line drawing images[oc95040d.gif] [oc95040a.gif] [oc95040e.gif] [oc95040b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}