 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias Instituto Oswaldo Cruz, Vol.90(2):221-227 mar./apr. 1995 Schistosomiasis Vaccine Development: Approaches and Prospects NR Bergquist UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR), WHO, Geneva, Switzerland
Code Number: OC95045
Size of Files:
Text: 34K
Graphics: Line Drawing(gif) - 17K
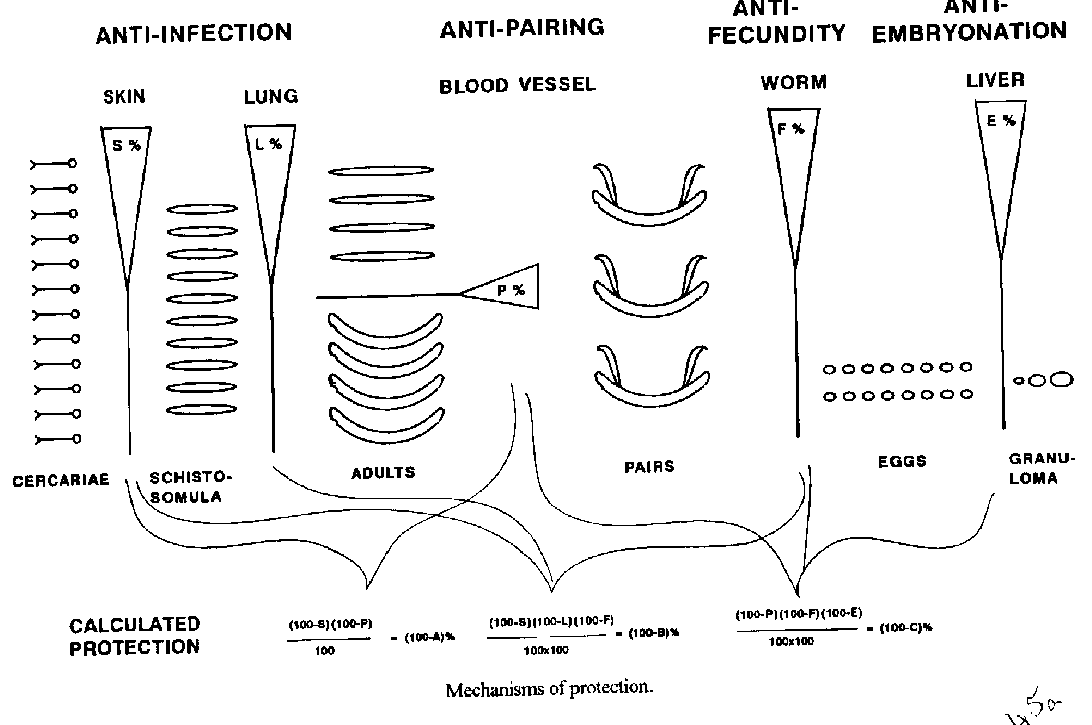
[TABLE AT END OF TEXT]Mounting evidence for acquired immunity to schistosomiasis in humans supports the case for immunological intervention. On the other hand, rapid reinfection poses a threat to younger age groups due to the slow maturation of natural resistance. However, rational approaches, based on advances in immunology and molecular biology, have substantially increased the odds of producing an effective vaccine. Since the parasite cannot replicate in the human host and serious morbidity generally occurs only after a relatively long period of heavy worm burden, complete protection against infection is not essential. The chances of success would increase if more than one of the various host/parasite interphases were targeted, for example reducing morbidity through decreased worm loads as well as through suppression of egg production. Several promising schistosome antigens have now reached an advanced phase of development and are currently undergoing independent confirmatory testing according to a standardized protocol. A few molecules are being contemplated for scaled- up production but, so far, only one has reached the stage of industrial manufacture and safety testing. Since schistosomiasis cannot realistically be controlled by a single approach, vaccination is envisaged to be implemented in conjunction with other means of control, notably chemotherapy. uKey words: antigen - cytokine - schistosomiasis - Schistosoma - immunity - protection - resistance - vaccine The ubiquity of reinfection after chemotherapy for schistosomiasis, particularly in young inhabitants of endemic areas, is a compelling reason for developing a long-term measure to complement drug treatment. Propelled by the accumulation of positive data on human acquired immunity from many geographical areas endemic for Schistosoma mansoni and/or S. haematobium (Hagan et al. 1991, Hagan & Abath 1992, Dessein et al. 1992, Butterworth et al. 1992), and by encouraging protection experiments in animal models with improved antigens, vaccine development must now be accepted as a realistic approach. Although gaps persist in our model of the development of pathology and resistance in schistosomiasis, a consolidated picture of the disease as the result of an interplay between opposing immunological mechanisms is emerging. Caught between the need to prevent further invasion of parasites and the necessity to modulate the granulomas which form around eggs trapped in host tissues, the immune system displays a bewildering activity of various effector cells and antibody isotypes. The main obstacle resides in the fact that parasites, in contrast to the great majority of other infectious agents, commonly strike a balance with their hosts, and one which does not exclude reinfection. In the case of schistosomiasis, however, several factors confer a comparative advantage, justifying the continued support of schistosomiasis vaccine development: (a) diverse protective mechanisms have been demonstrated in experimental animals; (b) naturally acquired partial immunity is the most plausible explanation for the observed reduced intensity of infection in adolescent humans and older age groups compared to children; (c) complete immunity is not required since the parasite does not replicate in the human host and the aim is confined to prevention of morbidity; (d) safe and effective drugs are avaible. Native antigens seem to provide superior protection but for reasons of safety and production parasite vaccines should be made from recombinant ones. Based on the assumption that candidate antigens are sufficiently cross-reactive to be effective against several species, it is prudent to work initially with S. mansoni whose life-cycle can easily be maintained in the laboratory. However, as it is only logical to expect that homologous antigens would produce improved protection, antigens of other schistosome species will eventually have to be developed. In fact, an impressive number of S. japonicum antigens has already been cloned and expressed (Waine et al. 1993). The possibility of finding new superior antigens cannot be ruled out but it makes sense to now move on to human trials with those molecules which are ready, not least because this would help elucidate how resistance against schistosomiasis develops in man, something which is still largely unknown. This paper emphasizes the advantages of a concerted approach to vaccine development geared at simultaneous interference with several stages of the parasite life-cycle. ANTIGEN COMPOSITION The biochemical nature of epitopes does not have implications only for immunogenicity. For example, carbohydrate antigens may augment the risk of inadvertently activating the granulo- matous reaction since they often cross-react with egg antigens (Simpson 1989). The contrast when comparing the relatively small granulomas of chronic schistosomiasis with the large, florid ones associated with the early stage of infection (Domingo & Warren 1968, von Lichtenberg 1987) and the fact that anti-carbohydrate antibodies eventually become down- regulated (Omer-Ali et al. 1989), accentuate the dualistic operation of the immune system. Antibodies which block the protective responses in S. mansoni (Butterworth et al. 1992) and S. haematobium (Hagan et al. 1991, Hagan et Abath 1992), probably constitute a related issue. VACCINE TARGETS The total number of cloned schistosome antigens, expressed in various microorganisms such as viruses, yeast and bacteria, including BCG, may soon reach 100 but only a few of them show genuine promise. The schistosomulum surface is the preferential antigen source but all stages of the parasite life-cycle in the vertebrate host may be targeted. Attempts to combine full-length protein antigens from the same stage have not been encouraging but the use of antigens from diverse stages might improve the possibilities for synergistic action by involving separate defence mechanisms. Table gives the current status of different approaches to obstruct the progress of the parasite, and Figure illustrates how consolidated levels of protection with several disparate antigens can be gauged. The arbitrarily chosen examples show that three antigens, each producing 25% protection, would together only allow about 40% of cercariae to develop to adult worms, whilst two antigens would require 40% protection each to surpass that. A brief discussion of potential components of a combination vaccine follows below. Skin penetration - Six cercarial antigens have been described. One is a 41 kDa antigen proposed for diagnosis of acute schistosomiasis (Hayunga et al. 1986) and the other five are proteases, measuring 25 kDa (Landsperger et al. 1982), 28 kDa (Marikovsky et al. 1990), 30 kDa (McKerrow et al. 1985), 41 kDa and 47 kDa (Chavez-Olortegui et al. 1992), respectively. These enzymes display diverse substrate specificities but have a strong immunogenicity in common, suggesting a possible role in vaccine development. Surprisingly, none of them have been seriously studied as possible candidate vaccine antigens. Larval development - The schistosomulum undergoes a phase of rapid growth from the time immediately after cercarial penetration until early maturity about three weeks later. The literature contains numerous references to surface antigens which range from 8 to >200 kDa and are often sha- red with other stages, most commonly with the adult worm and the egg (Simpson 1989, 1990). In general, the levels of protection displayed in permissive animals after immunization are not impressive and since it is well known that almost complete protection can be induced by cercariae attenuated by sub-lethal doses of radiation, the search for superior antigens continues. It should be noted, however, that this effect may not be due to the exposure of particular antigens per se but rather reflect improved antigen presentation due to a prolonged passage through lymph nodes and lungs. On the other hand, the significance of epitope configuration is emphasized by the demonstration of improved protection induced by some "new" antigens, including a 62 kDa myosin-like protein implicated in the irradiated cercariae model (Soisson et al. 1992) and Sm14 (Moser et al. 1991, Tendler et al. 1994) which have both shown protection in the 50-70% range in mice. Other effective, well-explored antigens associated with the schisto- somulum include paramyosin (Pearce et al. 1988), Sm23 (Reynolds et al. 1992, K”ster at al. 1993), triose-phosphatase isomerase (TPI) (Shoemaker et al. 1992, Reynolds et al. 1994) and, in particular, glutathione S-transferase (GST) (Balloul et al. 1987, Capron et al. 1992). Pairing - In vitro experiments have shown that the two sexes of schistosome do not attract each other before the fourth week of development, whilst older specimens orient themselves to the opposite sex to form egg-producing pairs (Eveland & Haseeb 1989). More recent research suggests that females exhibit a receptor capable of responding to a lipid- containing compound released from the male from the fourth week after infection (Haseeb & Eveland 1991). Identification of the receptor or the pheromone is a long-term endeavor but progress in this area could lead to a way of effectively blocking egg production, the major problem in schis- tosomiasis. Fecundity - Even after pairing has taken place, the production and release of eggs from the female schistosome may be successfully interfered with. Almost two decades ago, an anti-fecundity effect mediated by immune responses was reported in experimental studies on S. haematobium infection in baboons (Webbe et al. 1976) and later confirmed in cattle infected by S. bovis (Bushara et al. 1980). A fresh surge of interest in exploiting this potentially useful mechanism has brought these results to the forefront in vaccine discussions (Xu et al. 1991, Boulanger et al. 1991, Agnew et al. 1993). Interestingly, immunization of mice with S. mansoni GST indicates that this antigen not only protects against infection in the traditional sense but also reduces egg production by up to 85% (Xu et al. 1991, Boulanger et al. 1991). Granuloma formation - The early recognition of the phenomenon of granulomatous regulation (Domingo & Warren, 1968) opened up the prospects of anti-pathology vaccination, leading to the hypothesis of anti-embryonation (Garcia et al. 1982). This approach attempts to reduce immune-mediated pathology by halting egg maturation by vaccination with viable immature eggs, thereby curtailing the release of immunogenic compounds from the miracidium inside. Garcia et al. (1989) were able to show that S. japonicum eggs mature in lower numbers in chronically egg-sensitized mice and that sera from humans with chronic schistosomiasis are capable of reducing granuloma development. A beneficial effect on granuloma size after injection of irradiated schistosomula in baboons has been reported by Damian et al. (1984) and a similar but not statistically significant effect was noted after experimental vaccination with GST (Boulanger et al. 1991). However, as egg antigens are not involved in this case, the effect is probably not due to anti-embryonation. In an attempt to modulate the cell-mediated response to schistosomal antigens at the clonal level, T[H]1 cell clones directed against egg antigens have been raised and their targets identified (Chikunguwo et al. 1992, Stadecker, 1992, Flores-Villanueva et al. 1994). Further work along these lines may make it possible to induce anergy by blocking the events leading to the granulomatous reaction. CYTOKINE REGULATION The recently acquired ability to study the triggering of immune responses at the molecular level in vivo has revolutionized the study of host defence mechanisms. Resistance to infection in general is controlled by two pivotal sub-populations of T helper cells, T[H]1 and T[H]2, the former predominantly activating cell-mediated responses through the production of interferon-gamma (IFN-g), interleukin two (IL-2) and IL-12, whilst the latter is associated with IL-4 and IL-5 and the stimulation of antibodies including the immediate hypersensitivity and eosinophilia characteristic of helminth infection. The unravelling of these mechanisms in schistosomiasis opens the road to modulation of the immune response. For example, the injection of irradiated S. mansoni cercariae into mice results in a T[H]1 response correlated to resistance, whilst normal cercariae induces a T[H]2 response which is essentially without protection (Pearce et al. 1991, Grzych et al. 1991). The protective responses have been demonstrated to correlate with the activation of macrophage effector cells by IFN-g (James, 1992). Interestingly, the transfer of eggs to vaccinated mice incurs a switch from the T[H]1-dominated response to one mainly consisting of T[H]2 cells which seem to permit both infection and further egg deposition (Grzych et al. 1991). IL-4, IL-10 and transformation growth factor-beta (TGF-b) are potent down-regulating cytokines capable of inhibiting schistosomular destruction as shown by the fact that even suboptimal combinations of any two of these cytokines synergistically suppress parasite killing by IFN-g-activated macrophages (Oswald et al. 1992). For example, the supernatant from a line of spleen cells from a mouse infected with S. mansoni was reported to inhibit the growth of a culture of T[H]1 cells, which effect could be blocked by the addition of antibodies against IL-10 (Sher at al. 1991). In mice receiving macrophages isolated from egg granulomas IL-4 and IL-10 increased while IL-2 decreased to undectable levels (Flores-Villanueva et al. 1994). It is even possible that schistosomula use T[H]2 activation as a strategy to evade macrophage-mediated killing. On the other hand, anti-parasite IgG, IgE and IgA antibodies correlate with resistance to reinfection in drug-cured patients and eosinophils have been shown to mediate IgE and IgG2a antibody-dependent cell- mediated cytotoxicity against schistosomula in vitro (Capron et al. 1992, Dunne et al. 1992). IL-12 displays a strong T cell effect (Perussia et al. 1992) which could possibly be utilized for T[H]1 stimulation in future vaccines but since this cytokine also seems to down- regulate IgE responses (Kiniwa et al. 1992) there is again a balance to be struck. The growing awareness of the need to consider all aspects of the immune system, encourages the use of antigens expressing B cell epitopes as well as T cell epitopes. Most antigens proposed for vaccine studies contain both and some vaccine candidates consisting of synthetic peptides mimicking such epitopes have been produced and tested (Auriault et al. 1990, Reynolds et al. 1992, 1994). The granulomatous response to schistosome eggs appears to have a strong T[H]2 component, and its development is counteracted by treatment with anti IL-4 antibody (Chensue et al. 1992). The fibroblast-stimulating factor-1 (FsF-1), a fibrosis- stimulating cytokine produced by CD4+ cells residing in granulomas (Prakash & Wyler 1992) is another possible target for immunologic intervention. Finally, tumour necrosis factor (TNF)-a may play an important role in schistosome associated pathology since infection of the severe combined immunodeficiency (SCID) mouse followed by addition of the recombinant cytokine stimulates egg production and restores granuloma formation (Amiri et al. 1992). The role of the T[H]1/T[H]2 cell balance in humans with schistosomiasis remains unclear. On the one hand, a reverse balance of IFN-g/IL-4 persisting at least three months after cure, disturbed expressions of the ratio of CD3+ to CD8+ cells and a reversible decrease of IL-1 were reported in Brazilians infected with S. mansoni (Zwingenberger et al. 1989a,b, 1990). On the other, high levels of specific IgE antibodies correlate with post-treatment resistance in cohorts of Africans infected with S. mansoni or S. haematobium (Hagan et al. 1991, Dunne et al. 1992). CONCLUSIONS More emphasis on research into cercarial penetration of the host skin is warranted as an improved early impediment would reduce the number of developing schistosomula, thus facilitating implementation of preventive measures geared at the later stages. Combined antigens from different parasite stages should result in superior resistance and the less the relation between the responses elicited the better the chance of inducing mutually bolstering mechanisms. Rational antigen combination would permit also the utilization of antigens of modest protective value. It is, in this connection, interesting to note the natural combination of two, perhaps even three, separate effector mechanisms in the GST molecule. The risk of vaccine interference with the beneficial regulation of granuloma formation makes it necessary to avoid activating epitopes involved in this pathway. Even if it would be useful to reinforce natural down-regulation of the granuloma, this process might produce potentially harmful results if carried too far. For example, the toxic effects of leaking egg products could damage surrounding hepatocytes in the absence of any sequestering reactions. Stimulation of T cell anergy is an interesting approach but research is still at the basic stage. Antibody-mediated blocking of pairing of the two sexes is perhaps the most elegant way of avoiding pathological reactions due to schistosomiasis. Once the components of the chemical signaling system have been identified and characterized, generation of neutralizing antibodies against the pheromone, which probably avoids immune recognition due to its small size, would be straightforward. Alternatively, antibody-blocking of the receptor may be attempted. It is unfortunate that this potentially important area has so far attracted comparatively little attention and is consequently still far from applied research. There is obviously dependable support for the involvement of both cell-mediated and humoral mechanisms in schistosome infection but the complex regulation of specific immune responses clouds the picture. The coexistence of activated macrophages, different kinds of T-cells and antibodies, both effective and blocking ones, does not permit a simple solution. The enigma of how resistance is generated and maintained can only be penetrated further by initiating human Phase I/II vaccine trials. Safety is assured by the availability of efficacious drugs with few side effects. The degree of protection where a beneficial effect would be clearly observed can be estimated to be above 50% and is probably below 80%. Since intervention at this level would not arrest transmission, natural infection may be counted on to boost the artificially induced resistance. However, it might also be necessary to address the possibility of a slow build-up of the quantity of parasites to a significant level in subjects who are in regular contact with water containing infected snails. In addition, females tend to increase their individual egg production when their number in the host is reduced (Hayunga et al. 1985). For these reasons, chemotherapy will almost certainly have to be continued after vaccination in many areas, albeit with much longer intervals between treatments than called for in current schedules.
TABLE Defence Mechanisms and Intervention Targets Parasite activity Target Identified Cloned Tested ------------------------------------------------------- Cercarial invasion Antigens Some One None Larval growth Antigens Many Many Yes Worm pairing Pheromone None None None Receptor None None None Egg Production Antigens One Yes Yes Egg Maturation Miracidial None None None secretionREFERENCES Agnew AM, Murare HM, Navarrete Sandoval S, de Jong N, Krijer FW, Deelder AM, Doenhoff MJ 1992. The susceptibility of adult schistosomes to immune attrition. Mem Inst Oswaldo Cruz 87 Suppl. IV: 87-93. Amiri P, Locksley RM, Parslow TG, Sadick M, Rector E, Ritter D, McKerrow JH 1992. Tumour necrosis factor a restores granulomas and induces parasite egg-laying in schistosome- infected SCID mice. Nature 356: 604-607. Auriault C, Gras-Masse H, Pierce RJ, Butterworth AE, Wolowczuk I, Capron, M, Ouma JH, Balloul JM, Khalife J, Neyrinck JL 1990. Antibody response of Schistosoma mansoni infected human subjects to the recombinant P28 glutathione S- transferase and to synthetic peptides. J Clin Microbiol 28: 1918-1924. Balloul JM, Grzych JM, Pierce RJ, Capron A 1987. A purified 28 000 Dalton protein from Schistosoma mansoni adult worms protects rats and mice against experimental schistosomiasis. J Immunol 138: 3448-3453. Boulanger D, Reid GD, Sturrock RF, Wolowczuk I, Balloul JM, Grezel D, Pierce RJ, Otieno MF, Guerret S, Grimaud JA, Butterworth AE, Capron, A 1991. Immunization of mice and baboons with the recombinant Sm28GST affects both worm viability and fecundity after experimental infection with Schistosoma mansoni. Parasite Immunol 13: 473- 490. Bushara HO, Majid AA, Saad AM, Hussein MF, Taylor MG, Dargie JD, Marshall TF, Nelson GS 1980. Observations on cattle schistosomiasis in the Sudan, a study in comparative medicine. II. Experimental demonstration of naturally acquired resistance to Schistosoma bovis. Am J Trop Med Hyg 29: 442-451. Butterworth AE, Dunne DW, Fulford AJ, Thorne KJ, Gachuhi K, Ouma JH, Sturrock RF 1992. Human immunity to Schistosoma mansoni: observations on mechanisms and implications for control. Immunol Invest 21: 391-407. Capron A, Dessaint JP, Capron M, Pierce RJ 1992. Vaccine strategies against schistosomiasis. Mem Inst Oswaldo Cruz 87 Suppl IV: 19-27. Chavez-Olortegui C, Resende M, Tavares CA 1992. Purification and characterization of a 47 kDa protease from S. mansoni cercarial secretion. Parasitol 105: 211- 218. Chensue SW, Terebuh PD, Warmington KS, Hershey SD, Evanoff HL, Kunkel SL, Higashi GI 1992. Role of IL-4 and IFN-g in Schistosoma mansoni egg-induced hypersensitivity granuloma formation: orchestration, relative contribution, and relationship to macrophage function. J Immunol 148: 900-906. Chikunguwo SM, Harris TS, Brodeur PH, Harn DA, Stadecker MJ 1992. The cell-mediated response to schistosomal antigens at the clonal level: development and characterization of a panel of egg antigen-specific murine T cell clones. Eur J Immunol 22: 917-922. Damian RT, Roberts ML, Powell MR, Clark, JD, Lewis FA, Stirewalt, MA 1984. Schistosoma mansoni egg granuloma size reduction in challenged baboons after vaccination with irradiated cryopreserved schistosomula. Proc Natl Acad Sci USA 84: 3552-3536. Dessein A, Rihet P, Demeure C, Couisinier P, Bacellar O, Carvalho EM, Kohlstaedt S, Dessein H, Souza A, Prata A, Goudot V, Bourgois A, Abel L 1992. Factors controlling resistance to schistosomiasis in endemic areas. Med/sciences Synthese 8: 108-118. Domingo EO, Warren KS 1968. Endogenous desensitization. Changing host granulomatous response to schistosome eggs at different stages of infection with Schistosoma mansoni. Am J Pathol 52: 369-371. Dunne DW, Butterworth AE, Fulford AJ, Kariuki HC, Langley JG, Ouma JH, Capron A, Pierce RJ, Sturrock RF 1992. Immunity after treatment of human schistosomiasis: association between IgE antibodies to adult worm antigens and resistance to reinfection. Eur J Immunol 22: 1483-1494. Eveland LK, Haseeb MA 1989. Schistosoma mansoni: onset of chemoattraction in developing worms. Experientia 45: 309-310. Flores-Villanueva PO, Harris TS, Ricklan DE, Stadecker MJ 1994. Macrophages from schistosomal egg granuloma induce unresponsiveness in specific cloned Th-1 lympocytes in vitro and down-regulate schistosomal granulomatous disease in vivo. J Immunol 152: 1847-1855. Garcia EG, Mitchell GF 1985. Vaccination against immunopathological disease in schistosomiasis japonica. Asian Pac J Allergy Immunol 3: 140-142. Garcia EG, Rivera PT, Mitchell GF, Evardome RR, Almonte RE, Tiu WU 1989. Effects of induction of anti-embryonation immunity on liver granulomas, spleen weight and portal pressure in mice infected with S. japonicum. Acta Tropica 46: 93-99. Grzych JM, Pearce E, Cheever A, Caulada ZA, Caspar P, Heiny S, Lewis F, Sher A 1991. Egg deposition is the major stimulus for the production of Th2 cytokines in murine schistosomiasis mansoni. J Immunol 146: 1322-1327. Hagan P, Blumenthal UJ, Dunn D, Simpson AJ, Wilkins HA 1991. Human IgE, IgG4 and resistance to reinfection with Schistosoma haematobium. Nature 349: 243- 245. Hagan P, Abath FGC 1992. Recent advances in immunity to human schistosomiasis. Mem Inst Oswaldo Cruz 87 Suppl IV: 95-98. Haseeb MA, Eveland LK 1991. Schistosoma mansoni: a chemoattractive factor released by males and its receptor in females. Experientia 47: 970-974. Hayunga EG, Hillyer GV, Stek Jr M, Sumner MP, Duncan JF, Desanti C 1985. Attempted immunization of mice against Schistosoma mansoni by inoculation with purified glycoprotein antigens from adult worms. Proc Helminthol Soc Wash 52: 184-195 Hayunga EG, Mollegard I, Duncan JF, Sumner MP, Stek M Jr, Hunter, KW Jr 1986. Development of circulating antigen assay for rapid detection of acute schistosomiasis. Lancet 2(8509): 716-718 James SL 1992. Experimental models of immunization against schistosomes: lessons for vaccine development. Immunol Invest 21: 477-493. Kiniwa M, Gately M, Gubler U, Chizzonite R, Fargeas C, Delespesse G 1992. Recombinant interleukin-12 suppresses the synthesis of immunoglobulin E by interleukin-4 stimulated human lymphocytes. J Clin Invest 90: 262-266. Koster B, Hall MR, Strand M 1993. Schistosoma mansoni: immunoreactivity of human sera with the surface antigen Sm23. Exp Parasitol 77: 282-294. Landsperger WJ, Stirewalt MA, Dresden MH 1982. Purification and properties of a proteolytic enzyme from the cercariae of the human trematode parasite Schistosoma mansoni. Biochem J 201: 137-144. Marikovsky M, Arnon R, Fishelson Z 1990. Schistosoma mansoni: localization of the 28 kDa secreted protease in cercaria. Parasite Immunol 12: 389-401 McKerrow JH, Lindquist R, Werb Z, Pino-Heiss S 1985. Purification and characterization of an elastinolytic proteinase secreted by cercariae of Schistosoma mansoni. J Biol Chem 260: 3703-3707. Moser D, Tendler M, Griffiths G, Klinkert M-Q 1991. A 14-kDa S. mansoni polypeptide is homologous to a gene family of fatty acid binding proteins. J Biol Chem 266: 8447- 8454. Omer-Ali P, Mansour M, Woody JN, Smithers SR, Simpson AJG 1989. Antibody to carbohydrate and polypeptide epitopes on the surface of schistoso-mula of Schistosoma mansoni in Egyptian patients with acute and chronic schistosomiasis. Parasitol 98: 417-424. Oswald IP, Gazzinelli RT, Sher A, James SL 1992. IL-10 synergizes with IL-4 and transforming growth factor-beta to inhibit macrophage cytotoxic activity. J Immunol 148: 3578-3582. Pearce EJ, James SL, Hieny S, Lanar DE, Sher A 1988. Induction of protective immunity against Schistosoma mansoni by vaccination with schistosome paramyosin (Sm97), a nonsurface parasite antigen. Proc Natl Acad Sci USA 85: 5678-5682. Pearce EJ, Caspar P, Grzych JM, Lewis FA, Sher A 1991. Downregulation of Th1 cytokine production accompanies induction of Th2 responses by a parasitic helminth, Schistosoma mansoni. J Exp Med 173: 159-166. Perussia B, Chan SH, D Andrea A, Tsuji K, Santoli D, Pospisil M, Young D, Wolf SF, Trinchieri G 1992. Natural killer (NK) cell stimulatory factor or IL-12 has differential effects on the proliferation of TCR-alpha beta+, TCR-gamma delta+ T lymphocytes, and NK cells. J Immunol 149: 3495- 3502. Prakash S, Wyler DJ 1992. Fibroblast stimulation in schistosomiasis. XII. Identification of CD4+ lymphocytes within schistosomal egg granulomas as a source of an apparently novel fibroblast growth factor (FsF-1). J Immunol 148: 3583-3587. Reynolds SR, Shoemaker CB, Harn DA 1992. T and B cell epitope mapping of Sm23, an integral membrane protein of Schistosoma mansoni. J Immunol 149: 3995- 4001. Reynolds SR, Dahl CE, Harn D 1994. T and B epitope determination and analysis of multiple antigenic peptides for the S. mansoni experimental vaccine triose-phosphate isomerase. J Immunol 152: 193-200. Sher A, Fiorentino D, Caspar P, Pearce E, Mosmann T 1991. Production of L-10 by CD4+ T lymphocytes correlates with down- regulation of Th1 cytokine synthesis in helminth infection. J Immunol 147: 2713-2716. Shoemaker C, Gross A, Gebremichael A, Harn D 1992. cDNA cloning and functional expression of the Schistosoma mansoni protective antigen triose-phosphate isomerase. Proc Natl Acad Sci USA 89: 1842-1846. Simpson AJG Omer-Ali P, Meadows HM, Jeffs SA, Hagan P, Smithers SR 1989. Schistosoma mansoni surface antigens. Mem Inst Oswaldo Cruz 84 Suppl I: 179-187. Simpson AJG 1990. Schistosome surface antigens: developmental expression and immunological function. Parasitol Today 6: 40-45. Soisson LM, Masterson CP, Tom TD, McNally MT, Lowell GH, Strand M 1992. Induction of protective immunity in mice using a 62-kDa recombinant fragment of a Schistosoma mansoni surface antigen. J Immunol 149: 3612-3620. Stadecker MJ 1992. The role of T-cell anergy in the immunomodulation of schistosomiasis. Parasitol Today 8: 199-204 Tendler M, Vilar MM, Brito CA, Freire NMS, Katz N, Simpson A 1995. Vaccination Against Schistosomiasis and Fasciolasis with the New Recombinant Antigen Sm 14: Potential Basis of a Multi- Valent Anti-Helminth Vaccine? Men Inst Oswaldo Cruz 90: 255-256. Von Lichtenberg F 1987. Consequences of infection with schistosomes, p. 185-232. In D Rollinson & AJG Simpson (eds), The Biology of Schistosomes. Academic Press, London and New York. Waine GJ, Becker M, Kalinna BH, Yang W, Scott J, Liu X, Tiu W, McManus D 1993. Molecular vaccines against schistosomiasis: current status and the challenges ahead. Asian Pacific J Mol Biol Biotech 1: 26-35. Webbe G, James C, Nelson GS, Smithers SR, Terry RJ 1976. Acquired resistance to Schistosoma haematobium in the baboon (Papio anubis) after cercarial exposure and adult worm transplantation. Ann Trop Med Parasitol 70: 411-424. Xu Chuan-Bo, Verwaerde C, Grzych J-M, Fontaine J, Capron A 1991. A monoclonal antibody blocking the Schistosoma mansoni 28-kDa glutathione S-transferase activity reduces female worm fecundity and egg viability. Eur J Immunol 21: 1801-1807. Zwingenberger K, Irschick E, Siqueira-Vergetti JG, Correia- Dacal AR, Janssen-Rosseck R, Bienzle U, Huber C, Feldmeier H 1989a. Release of interleukin 2 and gamma interferon by peripheral mononuclear cells in human Schistosoma mansoni infection normalizes after chemotherapy. Scand J Immunol 30: 463-471. Zwingenberger K, Harms G, Vergetti de Siqueira JG, Correia- Dacal AR, Janssen-Rosseck R, Bienzle U, Feldmeier H 1989b. T- cell phenotype alterations in hepatosplenic schistosomiasis mansoni normalize after chemotherapy. Immunobiol 179: 342-352. Zwingenberger K, Richter J, Taupitz S, Siqueira-Vergetti JG, Correia-Dacal AR 1990. Altered generation of interleukin 1 in chronic human schistosomiasis mansoni. Scand J Immunol 31: 729-736. Copyright 1995 Fundacao Oswaldo Cruz (Fiocruz)
The following images related to this document are available:Line drawing images[oc95045a.gif] |
| |||||||||

{kind=link}