 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias Instituto Oswaldo Cruz, Vol. 90(2):249-253 mar./apr. 1995 Comparison of Purified 12 kDa and Recombinant 15 kDa Fasciola hepatica Antigens Related to a Schistosoma mansoni Fatty Acid Binding Protein George V Hillyer Laboratory of Parasite Immunology and Pathology, Department of Pathology and Laboratory Medicine, UPR School of Medicine, GPO Box 365067, San Juan, Puerto Rico 00936-5067
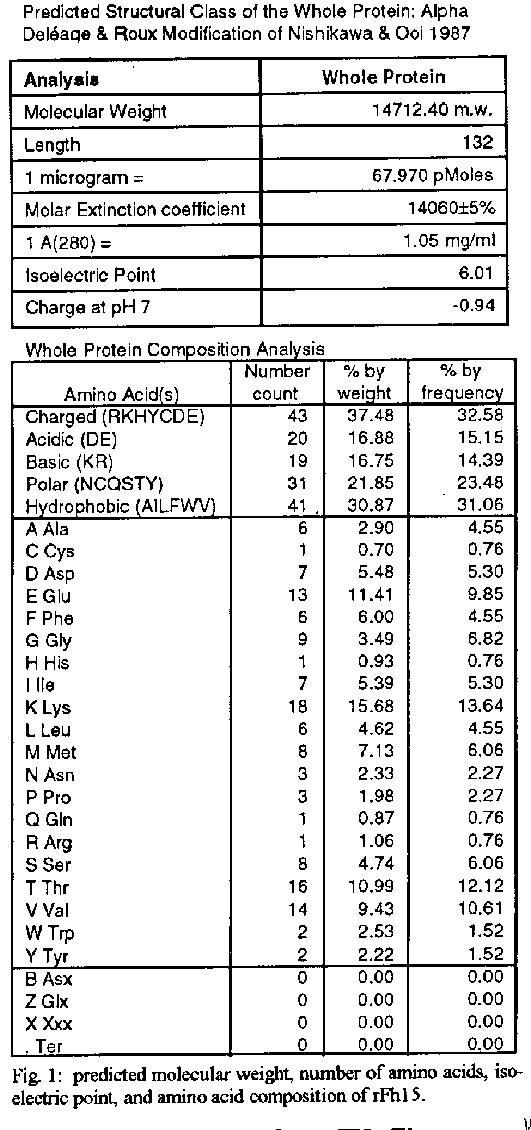
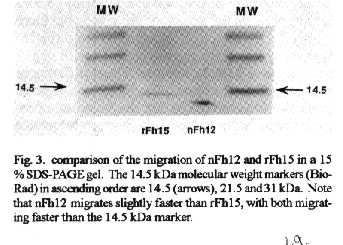
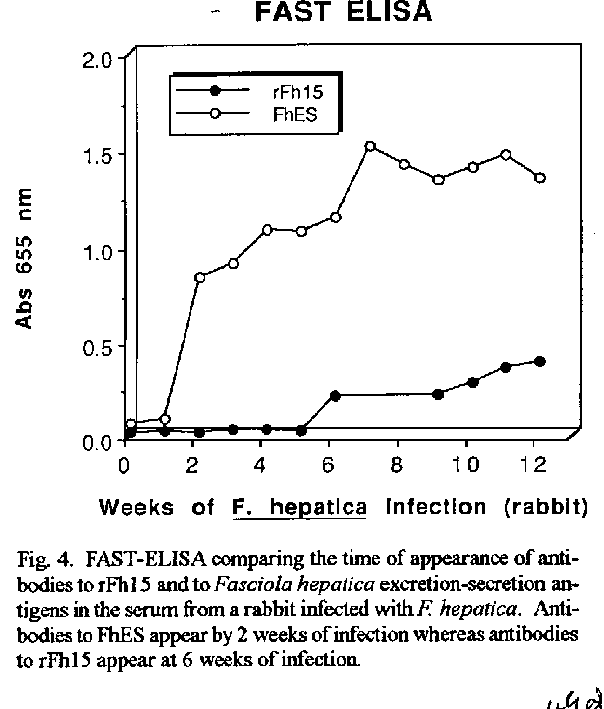
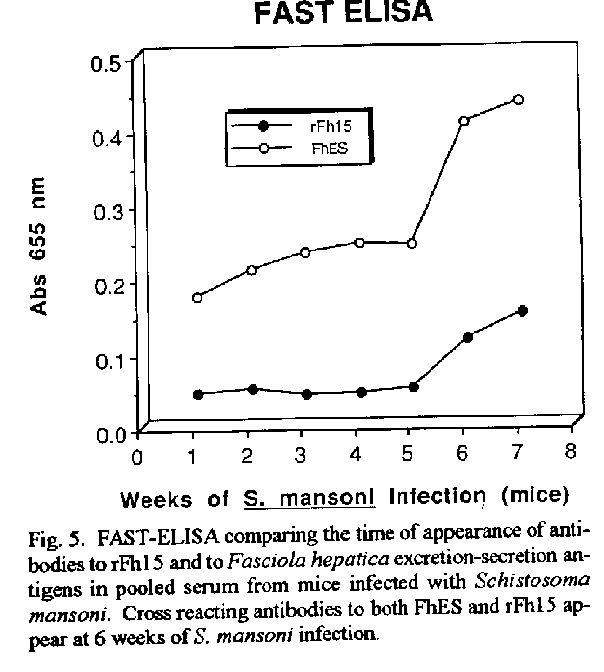
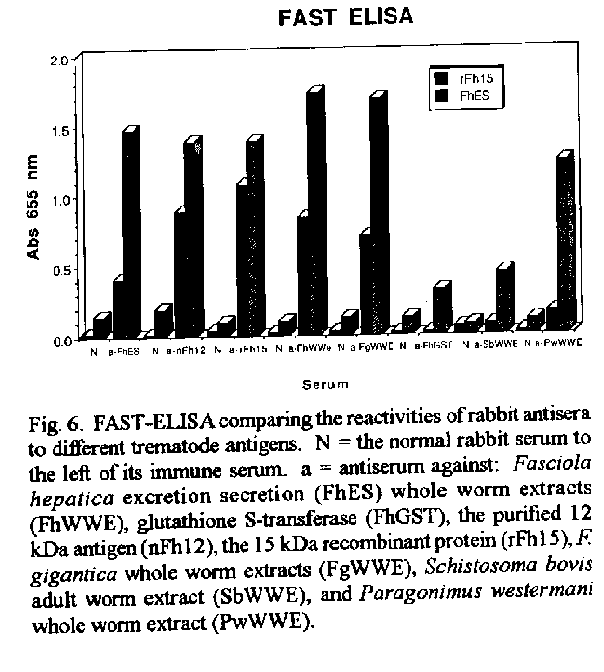
Vaccines in schistosomiasis using homologous antigens have been studied extensively in experimentally infected mammalian hosts. Vaccines using heterologous antigens have received comparatively less attention. This review summarizes recent work on a heterologous 12 kDa Fasciola hepatica antigenic polypeptide which cross reacts with Schistosoma mansoni. A cDNA has been cloned and sequenced, and the predicted amino acid sequence of the recombinant protein has been shown to have significant (44 %) identity with a 14 kDa S. mansoni fatty acid binding protein. Thus in the parasitic trematodes fatty acid binding proteins may be potential vaccine candidates. The F. hepatica recombinant protein has been overexpressed and purified and denoted rFh15. Preliminary studies show that rFh15 migrates more slowly (i.e. may be slightly larger) than nFh12 on SDS-PAGE and has a predicted pI of 6.01 vs. observed pI of 5.45. Mice infected with F. hepatica develop antibodies to nFh12 by 2 weeks of infection vs. 6 weeks of infection to rFh15; on the other hand, mice with schistosomiasis mansoni develop antibodies to both nFh12 and rFh15 by 6 weeks of infection. Both the F. hepatica and S. mansoni cross-reactive antigens may be cross-protective antigens with the protection inducing capability against both species. Key words: Fasciola hepatica - Schistosoma mansoni - heterologous resistance - Fasciola/Schistosoma cross-reactivity - fatty acid binding protein VACCINES IN SCHISTOSOMIASIS Several vaccine candidates have been identified in Schistosoma mansoni directed against the schistosomulum as well as against other life cycle stages. Some of the more promising antigens have now reached a more advanced stage of development including in the case of glutathione S-transferase, the stage of industrial manufacture and safety testing (Capron 1992, Bergquist et al. 1994). Another approach has been to study closely related cross-reacting antigens from another trematode, Fasciola hepatica. One 12 kDa antigen has been shown to induce in outbred albino mice significant levels of resistance to challenge infection with S. mansoni (Hillyer et al. 1988a, 1990). This Fasciola antigen is most probably related to a recently described immunoprotective fatty acid binding protein from S. mansoni. This review covers recent studies with this heterologous antigen from F. hepatica. HETEROLOGOUS RESISTANCE IN SCHISTOSOMIASIS AND FASCIOLIASIS Many studies have demonstrated that infection with one species of parasite can induce significant reduction in infection with another parasite (heterologous immunity or resistance). This reduction is manifested by reduced parasite burdens relative to the challenge infection, reduced egg laying (anti-fecundity), reduced pathology, or a combination of these. Studies on heterologous resistance in relation to schistosomes and Fasciola were summarized at the 2nd International Symposium on Schistosomiasis (Hillyer 1987). Using the murine model investigators have shown that infection with F. hepatica induces high levels of resistance to challenge infection with S. mansoni and vice versa (Hillyer 1987, Christensen et al. 1987, Hillyer et al. 1988b). This heterologous resistance has an immunologic basis since the Fasciola antigen(s) conferring resistance cross-reacts with S. mansoni (Hillyer et al. 1988a,b). RESISTANCE TO SCHISTOSOMES USING DEFINED, HETEROLOGOUS FASCIOLA ANTIGENS A F. hepatica 12 kDa (nFh12) polypeptide purified from adult worm extracts induces in mice significant levels of resistance to challenge infection with this parasite. In four separate experiments, outbred albino NIH (GP) mice immunized with nFh12 and challenged percutaneously with S. mansoni cercariae developed 47, 52, 60, and 77 % fewer schistosome worms than controls (Hillyer et al. 1988a, 1990). The term nFh12 is used to define the purified native polypeptide. On SDS-PAGE this polypeptide migrates faster than the 14 kDa MW marker and its migration is calculated to be ca. 12 kDa. It is a potent immunogen which must be expressed early after the transformation of the metacercaria cyst stage to the juvenile stage because different animal species (mice, calves, rabbits) infected with F. hepatica develop antibodies to Fh12 by 2 weeks of infection (Hillyer et al. 1988b). It is a cross-reactive antigen because mice infected with S. mansoni develop antibodies to Fh12 by 5 weeks of infection. MOLECULAR CLONING OF A cDNA EXPRESSING RECOMBINANT nFh12 (rFh15) A lambda gt11 F. hepatica cDNA library was constructed from poly(A)+ RNA extracted from adult worms. The unamplified library had a titer of 2.2 X 104 plaque-forming units/ml and 99 % recombinants. Primary screening of 300,000 phage plaques from the amplified library with an anti-Fh12 antiserum resulted in 33 positive clones. After plaque purification, 17 recombinant phages were selected for further analysis. One cDNA was sequenced and the predicted amino acid sequence revealed an open reading frame encoding a 132 amino acid protein with a predicted molecular weight of 14,700 Da (Rodriguez-Perez et al. 1992). This recombinant antigen was denoted rFh15. It has a predicted molecular weight of 14.7 kDa, 132 amino acids, and an isoelectric point of 6.01. These parameters and others including the predicted amino acid composition are summarized in Fig. 1. Importantly, the predicted amino acid sequence of rFh15 has significant identity (44 %) to a 14.8 kDa S. mansoni fatty acid binding protein (Moser et al. 1991). Because of the similarity of the Fasciola/Schistosoma protection inducing, cross reactive antigen (rFh15) with Sm14, our prediction has been that the schistosome homologue must also be a protection inducing molecule against S. mansoni and, possibly, F. hepatica (Rodriguez-Perez et al. 1992). CLONING, OVEREXPRESSION AND PURIFICATION OF rFh15 The Fh15 coding sequences were amplified by polymerase chain reaction, and the purified, amplified Fh15 DNA and the vector pGEX2TK were digested with BAM HI. The gel purified, digested PCR product was ligated into the BAM HI digested and dephosphorilated pGEX2TK. The overexpressed S. japonicum glutathione S-transferase (SjGST)-rFh15 fusion protein was affinity purified using glutathione agarose by competitive elution with excess reduced glutathione (Smith & Johnson 1988). Pure rFh15 is obtained after thrombin cleavage of the fusion protein and subsequent purification using a second glutathione agarose column (Hillyer, Laxer, Soler de Galanes, Garcia-Blanco in prep.). Major details of this purification procedure are found in Fig.2. PRELIMINARY COMPARISONS OF nFh12 AND rFh15 When pure nFh12 and rFh15 are compared on a 15 % SDS-PAGE gel, both migrate faster than the 14 kDa molecular weight standard, but nFh12 always migrates slightly faster than rFh15 (Fig. 3). This suggests that rFh15 may have a slightly higher MW than nFh12. The predicted pI of rFh15 is 6.0; the observed pI using a BioRad Rotofor with 4-6 pH ampholytes is 5.45 (Hillyer, Laxer, Soler de Galanes, Garcia-Blanco in prep.). Direct comparisons of the sequences of both proteins will resolve these differences and are in progress. Other results on the time of appearance of antibodies during infections support differences between nFh12 and rFh15. For example, mice, calves, and rabbits develop antibodies to F. hepatica excretion-secretion (FhES) antigens and to nFh12 by two weeks of infections (Hillyer et al. 1988b, Hillyer & Soler de Galanes 1991). In contrast, in rabbits with fascioliasis antibodies to rFh15 appear much later, i.e. by the 6th week of infection (Fig. 4). Mice infected with S. mansoni develop antibodies which are cross reactive with FhES and nFh12 (Hillyer et al. 1988b, Hillyer & Soler de Galanes 1991), as well as to rFh15 (Fig. 5) by the 6th week of infection. Thus the differences in timing of appearance of antibodies between nFh12 and rFh15 are in the fascioliasis, but not schistosomiasis, infections. It was also of interest to determine the reactivity of different rabbit antisera with purified rFh15. These are seen in Fig. 6. Antisera to F. hepatica excretion-secretion, whole worm antigens, and F. gigantica whole worm antigens, clearly react with rFh15, as do antisera to nFh12 and rFh15. Antisera to F. hepatica glutathione S-transferase did not react with rFh15. Antisera to S. bovis and to Paragonimus westermani had lesser reactivity, but cross reactivity was clearly evident. These last two are of special interest with regard to heterologous resistance because Hillyer and Serrano (1983) demonstrated that mice immunized with P. westermani adult worm extracts developed resistance to challenge infection with S. mansoni. More recently, Rodriguez-Osorio et al. (1993) showed that sheep infected with F. hepatica for 10 weeks developed 87 % less S. bovis worms than controls. This is similar to the finding that sheep infected with S. mansoni for 10 weeks acquired resistance to challenge infection with F. hepatica (Haroun & Hillyer 1988). Heterologous resistance has also been demonstrated by Yagi et al. (1986) who found that F. gigantica infections protected cattle against challenge infection with S. bovis. Because the homologues of rFh15 (or nFh12) may be involved in protective heterologous immunity, further studies are required to demonstrate that they are also present in the heterologous antigen preparations used to prepare the antisera described above. It should be noted that rabbit antisera against Fh12 differ in their reactivities depending on the method of isolation of antigen. For example, an anti-Fh12 antiserum developed by immunization with Fh12 excised from SDS-PAGE gels when reacted with crude F. hepatica adult worm extracts (FhWWE) will yield a single band on SDS-PAGE at the 12 kDa position by Western Blot. This same antiserum will fail to react with FhWWE in FAST-ELISA. In contrast, anti-Fh12 developed by immunization with Fh12 purified by a combination of gel filtration and isoelectric focusing will react with SDS-PAGE fractionated FhWWE in Western Blot and also by FAST-ELISA (data not shown). These results suggest that conformational epitopes must be taken into account when preparing antisera to antigens isolated in different manners. FATTY ACID BINDING PROTEINS AS VACCINES IN FASCIOLIASIS AND SCHISTOSOMIASIS Two fatty acid binding proteins have been described from adult worm parasitic platyhelminthes: a protein with 133 amino acids from S. mansoni denoted Sm14 (Moser et al. 1991), and a 15.5 kDa protein from Echinococcus granulosus denoted Eg15 (Esteves et al. 1993). Amino acid sequence alignment and comparison of rFh15 with Sm14 shows 44 % identity among them and thus suggests that the F. hepatica antigen may also be a fatty acid binding protein. Moreover, extensive studies by M Tendler and collaborators in Brazil have shown that mice and rabbits immunized with recombinant Sm14 develop significantly less S. mansoni worms after challenge than controls. Similarly, mice immunized with recombinant Sm14 develop high levels of resistance to challenge with F. hepatica (M Tendler, Intl. Symp. on Schistosomiasis, Rio de Janeiro, Dec. 1993). Moreover, our own studies have shown that nFh12 protects mice against challenge with S. mansoni (Hillyer et al. 1988a, 1990). Are these three FABPs common protective antigens? A comparison of the deduced amino acid sequences of all three are shown in Fig.7. It is clear that rFh15 is more closely related to Sm14 than to Eg15 with 43.9 % identity between the trematode sequences; but there is only 23.5 % identity between rFh15 and rEg15. Moreover, direct comparison of Sm14 with rEg15 only shows a 33.8 % identity. These differences may be sufficient to preclude rEg15 as a vaccine candidate for cestodes as are the trematode proteins if immunity is based on primary sequence alone. But if protection is a function of blocking the biological activity of the fatty acid binding protein, then the sequence divergence may be less important than the function of the protein. For example, Sacchettini and Gordon (1993) have shown that comparisons of numerous mammalian and avian fatty acid binding proteins reveal similar conformations even though multiple sequence alignments reveal that they have as little as 20 % amino acid sequence identity. Thus the protection inducing activity of Eg15 needs to be tested directly. Homologous recombinant antigens which are potential vaccines against schistosomes have been tested extensively (Capron 1992, Bergquist et al. 1994). The trematode fatty acid binding proteins may soon join this list of vaccine candidates. REFERENCES Bergquist NR, Hall BF, James S 1994. Schistosomiasis vaccine development: Translating basic research into practical results. The Immunologist in press. Capron A 1992. Immunity to schistosomes. Curr Op Immunol 4: 419-424. Christensen NO, Nansen P, Fagbemi BO, Monrad J 1987. Heterologous antagonistic and synergistic interactions between helminths and between helminths and protozoans in concurrent experimental infection of mammalian hosts. Parasitol Res 73: 387-410. Esteves A, Dallagiovanna B, Ehrlich R 1993. A developmentally regulated gene of Echinococcus granulosus codes for a 15.5-kilodalton polypeptide related to fatty acid binding proteins. Mol Biochem Parasitol 58: 215-222. Haroun ETM, Hillyer GV 1988. Cross-resistance between Schistosoma mansoni and Fasciola heptica in sheep. J Parasitol 74: 790-795. Hillyer GV 1987. Heterologous resistance in schistosomiasis. Mem Inst Oswaldo Cruz 82 (Suppl. IV): 171-174. Hillyer GV, Garcia Rosa MI, Alicea H, Hernandez A 1988a. Successful vaccination against murine Schistosoma mansoni infection with a purified 12 KD Fasciola hepatica cross-reactive antigen. Am J Trop Med Hyg 38: 103-110. Hillyer GV, Garcia Rosa MI, Soler de Galanes M 1990. Identification of Fasciola hepatica molecules with immunodiagnostic and immunoprophylactic potential, p. 239-259. In R Ehrlich, A Nieto, L Yarzabal, Basic Research in Helminthiases. Ediciones LOGOS, Montevideo, Uruguay. Hillyer GV, Serrano A 1983. The antigens of Paragonimus westermani, Schistosoma mansoni, and Fasciola hepatica adult worms. Evidence for the presence of cross-reactive antigens and for cross-protection to Schistosoma mansoni infections using antigens of Paragonimus westermani. Am J Trop Med Hyg 32: 350-358. Hillyer GV, Soler de Galanes M 1991. Initial feasibility studies of the FAST-ELISA for the immunodiagnosis of fascioliasis. J Parasitol 77: 362-365. Hillyer GV, Soler de Galanes M, Garcia Rosa MI, Montealegre F 1988b. Acquired immunity in schistosomiasis with purified Fasciola hepatica cross-reactive antigens. Vet Parasitol 29: 265-280. Moser D, Tendler M, Griffiths G, Klinkert M 1991. A 14-kDa Schistosoma mansoni polypeptide is homologous to a gene family of fatty acid binding proteins. J Biol Chem 266: 8847-8454. Rodriguez-Osorio M, Gomez-Garcia V, Rojas-Gonzalez J, Ramajo-Martin V, Manga-Gonzalez MY, Gonzalez-Lanza C 1993. Resistance to Schistosoma bovis in sheep induced by an experimental Fasciola hepatica infection. J Parasitol 79: 223-225. Rodriguez-Perez J, Rodriguez-Medina JR, Garcia-Blanco MA, Hillyer GV 1992. Fasciola hepatica: Molecular cloning, nucleotide sequence, and expression of a gene encoding a polypeptide homologous to a Schistosoma mansoni fatty acid binding protein. Exp Parasitol 74: 400-407. Sacchettini JC, Gordon JI 1993. Rat intestinal fatty acid binding protein. A model system for analyzing the forces that can bind fatty acids to proteins. J Biol Chem 268: 18399-18402. Smith DB, Johnson KS 1988. Single-step purification of polypeptides expresed in Escherichia coli as fusions with glutathione S-transferase. Gene 67: 31-40. Yagi AI, Younis SA, Haroun EM, Gameel AA, Bushara HO, Taylor MG 1986. Studies on heterologous resistance between Schistosoma bovis and Fasciola gigantica in Sudanese cattle. J Helminthol 60: 55-99. Copyright 1995 Fundacao Oswaldo Cruz (Fiocruz)
The following images related to this document are available:Halftone images[oc95049c.gif]Photo images[oc95049c.jpg]Line drawing images[oc95049f.gif] [oc95049g.gif] [oc95049d.gif] [oc95049a.gif] [oc95049e.gif] [oc95049b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}