 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias Instituto Oswaldo Cruz, Vol. 90(2):261-269 mar./apr. 1995 Current Advances on the Study of Snail-Snail Interactions, with Special Emphasis on Competition Process Jose Rabelo de Freitas*, Mairy Barbosa Loureiro dos Santos Departamento de Biologia Geral, ICB, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627, 31270-901 Belo Horizonte, MG, Brasil
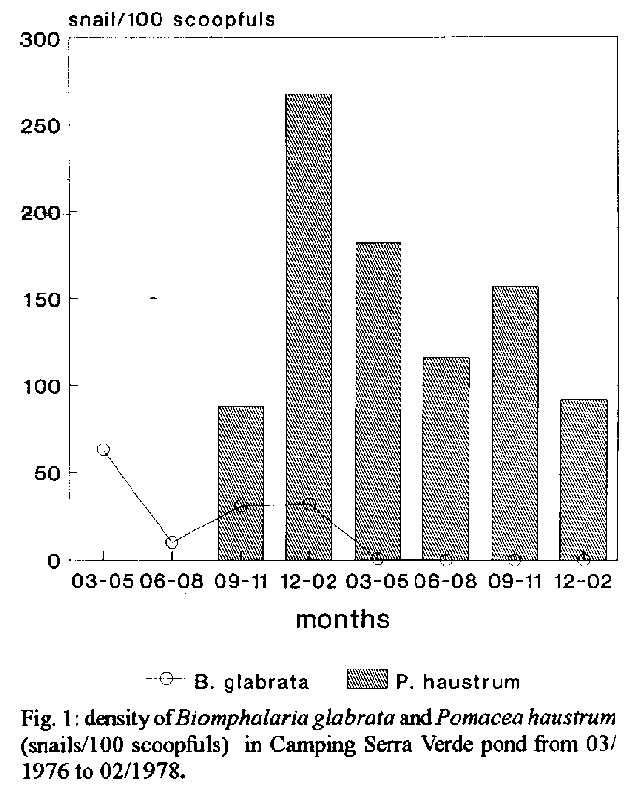
[TABLES AT END OF TEXT]Field work research on population dynamic of snails from the regions of Belo Horizonte and Lagoa Santa give much information about interactions among two or more species of mollusks: Pomacea haustrum, Biomphalaria glabrata, B. tenagophila, B. straminea and Melanoides tuberculata. Data ranging from two years to several decades ago suggest that the Pampulha reservoir is like a cemetery of B. glabrata and B. straminea, species that coexist for more than 14 years in a small part of a stream, whereas only B.glabrata lives in all the streams of the basin. In the last ten to twenty years B. tenagophila has coexisted with P. haustrum and M. tuberculata in the Serra Verde ponds and in the Pampulha dam. However these species have not settled in any of the brooks, except temporarily. The data suggest that the kind of biotope and the habitat conditions are decisive factors for the permanence of each species in its preferencial biotope. B. glabrata, natural from streams and riverheads, quickly disappears from the reservoirs and ponds where it coexists with other spcies for a short time, independently of the competitive process.Competition needs to be better studied, since in Central America and Caribean islands this kind of study has favored the biological control of planorbid species. Key words: biological control - snail competition - Biomphalaria tenagophila - Melanoides tuberculata To define competition is not simple nor are the mechanisms involved very obvious (McArthur 1972). All definitions of competition emphasize the affinity among species and the availability of resources. Scarcity makes each individual spend more energy in search of food. A reduced availabitity of resources results in a decrease of growth, reproduction and survival of the competitors. According to Tillman (1982), a resource is any substance or factor which can lead to increased growth rates. Food, space, light, water, soil, nutrients, etc. are generally considered resources. Each species may coexist on the condition that it varies somehow from one to the other possible competitors in the use of resources. Only the ones which have already adapted to different diets may coexist. That is why similar species are seldom found in the same biotope. However, species may coexist without competition when there are marked phylogenetic differences among them, since each species has its own "niche" - word difficult to fit in any concept (McArtuhr 1972). Among the American planorbids there seems to be no register of two similar species of hosts of Schistosoma, coexisting in the same primitive habitat. The occasional and transitory coexistence between the similar species Biomphalaria glabrata and B. tenagophila has been detected in a few secondary biotopes (Magalh„es 1966, Paraense 1970, Kavazoe et al. 1980). The rare examples of cohabitation generally refer to species of very little affinity such as between B. glabrata and B. straminea. In this case, the period of coexistence has sometimes been short due to exclusion of one of the species (Barbosa 1973) and in several occasions the coexistence is long and apparently without perspective of exclusion. Aspects related to competitive interactions between mollusks will be discussed and analysed, with emphasis on Pomacea haustrum, Melanoides tuberculata and Biomphalaria species in the surrounding cities of Belo Horizonte and Lagoa Santa. Data on competitive interaction among Brazilian snails will be compared with those mainly observed in Central America. There, many studies have been carried out with success of pilids and thiarids in the biological control of mollusk species host of Schistosoma mansoni. COMPETITIVE INTERACTIONS BETWEEN PILID AND PLANORBID SPECIES IN FIELD CONDITIONS Marisa cornuarietis was the most studied species used as a competitor of planorbids and bulinids until 1968. This snail is a competitive feeder and incidental predator on the eggs and young B. grablata (Sturrock 1974). The introduction of M. cornuarietis in almost a hundred ponds and irrigation canals, harboring a dense population of B. glabrata, caused the displacement of this snail in Puerto Rico, in a short time (Ruiz 1968, Ruiz-Tiben et al. 1969). The introduction of M. cornuarietis in Guadalupe decreased B. grablata population when it was associated with Pistia stratiotis (Pointier et al. 1991). Ferguson (1972) and Prentice (1983) reported the success of M. cornuaretis in some kind of habitats like ponds in Porto Rico and Africa. Nevertheless, in Tanzania, M. cornuarietis decreased population of four snail species but was not successful when colonizing habitats like streams. Nguma et al. (1982) reported the difficulty to get successful introduction of competitors. In Brazil few studies at field and laboratory have been done using P. haustrum as acompetitor (Guimar es l978, Andrade & Carvalho 1979). However, many studies on population dynamics of planorbids in Belo Horizonte and Lagoa Santa bring many information about the possible interactions among these snails and P. haustrum. Field experiments with the aim of controlling B. glabrata population by the pilid P. haustrum in Brazil, was carried out in Baldim and Calciolƒndia, Minas Gerais and at Ourinhos, S o Paulo. P. haustrum had been introduced into ditches, lakes and streams at Baldim, replaced B. grablata in many biotopes, included streams, and was predominant in others (Andrade & Carvalho 1979). However, despite its density reduction, B. glabrata was present in four biotopes out of seven, together with P. haustrum, 10 years after its introduction. In Calciolƒndia P. haustrum placed into five biotopes of B. grablata, within little time, reached densities from 200 to 600 individuals per month in ditches, and rates about 10 times higher, approximately, in the lake and brooks. The planorbid was no longer present in the biotopes, after four years coexistence with the pilid (Guimar es 1978). The fluctuations in density of the two species coexisting together, did not differ very much from those concerning the control of the experiment. In both situations (test and control) before and after the introduction of P. haustrum, B. grablata sometimes disappeared for several consecutive months, mainly in April and June, exactly when the collections were interrupted in the last two years. Because of this, we can not say that the exclusion really occurred. In spite of the coexistence of P. haustrum with B. glabrata and B. tenagophila for five years with rather high densities at Ourinhos, SP, it could not be verified the influence of the pilid on planorbids (Kavazoe et al. 1980). Seldom was P. haustrum found in Pampulha brooks and springs, habitats of B. grablata, except on their watery and swampy borders. Usually not only P. haustrum but also M. cornuarietis apparently do not adapt well into streams, but only in ponds, lakes and reservoirs, all still waters, as well in Santa Lucia banana drains (Ferguson & Tiben 1971, Sturrock 1974, Jordan et al. 1980). INTERACTIONS BETWEEN P. HAUSTRUM AND B. GLABRATA IN PAMPULHA RESERVOIR, SERRA VERDE PONDS AND LAGOA SANTA LAKE P. haustrum and B. grablata in Pampulha reservoir - There was a population explosion of B. grablata, which occupied in 1938 the whole border of the newly built Pampulha reservoir. Few years later, there was a drastic reduction of its density and area of occurrence, and that population almost disappeared. In the early 1950's the planorbid appeared only in the mouth of the streams, while P. haustrum was largely present, occupying all the border and appeared at the seventy-two collection sites (Martins & Falc o 1953, Andrade 1959). During many years, the ratio between P. haustrum and B. grablata could be estimated in 100 to 200 pilids for each single planorbid (Guimar es 1978). In the following years, B. grablata showed averages inferior to 1,3 specimen for each 100 scoopfuls/year (Carvalho et al. 1985) and during several months the density was 0 (zero). At depth of 0.50 to 2.0 m the pilid population was much lower than on the Pampulha borders and B. grablata was never collected at these depths. From 1990 onward all the collections have shown absence of planorbids. The Pampulha dam, like the ponds, look like a cemetery of B. grablata. P. haustrum and B. glabrata in Camping Serra Verde Pond - In the newly built Camping Serra Verde pond a dense population of B. grablata, coming from the springs and swampy waters of the brooks, was developed in the pond and from March 1976 showed a stable and gross population in the first year, a sharp fall of its density in the rainy season and disapperance in the second year. P. haustrum had its density treble during the three months following the invasion, in September, 1976 and kept high average densities, and peaks of 88 to 268 pilids/100 scoopfuls each three months. The proportion was generally from three to eight P. haustrum for each B. grablata (Fig.1) The disappearance of B. grablata from the Camping pond could be ascribed to the presence of P. haustrum bearing great numerical superiority. However, the two species lived together in the pond only four months. P.haustrum and B.glabrata in Hippodrome Serra Verde Pond - In the Hippodrome pond, the average density of B. grablata was superior to 25 snails for each 100 ladlefuls, per collection station, per month, but it reached monthly peaks of over 100 snails. Besides great fluctuations of population, B. glabrata showed very low densities, alternating with nonexistent ones (zero), for several successive months, mainly during the rainy season. The first specimens of P. haustrum appeared on the second month after the two last negative collections of B. glabrata. Therefore, the two species did not coexist during the monitoring period. Few years later, B. tenagophila invaded the two Serra Verde ponds, when B. grablata had already disappeared from these small dams, but it is possible that it lives in the headwater of the spring streams what has occurred with frequency. In these ponds we may found the coexistence of B. tenagophila and P. haustrum for many years. P. haustrum and B. glabrata in Lagoa Santa lake - B. glabrata present in the borders of this lake for some years in the l950, (Paraense & Santos l953) was eliminated few years later without any new register in this lagoon. It can be supposed that the disappearance of B. glabrata has nothing to do with the competitive action of P. haustrum or B. straminea. This asssumption is based on the fact that B. glabrata, present in springs and brooklets downstream, whose waters flow from the lake, could have invaded it by rheotaxis (Jurberg et al. 1988) and tried to settle there. This could have occurred other times, mainly during seven years (1979-1986) when in the lake, there was neither P. haustrum nor B. straminea nor any other possible competitive species of mollusk, including M. tuberculata, absent yet. INTERACTIONS BETWEEN P. HAUSTRUM AND B. STRAMINEA IN LAGOA SANTA AND LAGOINHA LAKES In Lagoa Santa lake - P. haustrum showed the highest densities in the shallowest area covered with rush (Eleocharis sp.), while B. straminea lived mainly in the region of Chara sp. at a depth ranging from 2,0 to 5,0 m. In 1978, 92% of this last species were collected in this area. The complete disappearance of the whole rush community that surrounded the edge of the lake, owing to the raising of its water level in 1979, led to the extinction of the aquatic macrophytes and all the mollusks including P. haustrum and B. straminea coexisting in the lake for more than 20 years, associated with the rush and Chara sp. stands. The elimination of the macrophytes Pistia stratiotis in Guadalupe, and also, as a consequence, the elimination of B. grablata, was done through the pilid snails (Pointier et al. 1991) not as a result of the desctruction of the habitats of all the mollusks as it occurred in Lagoa Santa. These environmental control measures and others like drainage of springs, all of them including elimination of macrophytes, have been greatly successful against B. grablata in Itabira, Minas Gerais. However, it should be pointed out that the study on planorbid-macrophyte interaction has only recently become important (Pieri & Thomas 1987, Thomas 1987, Santos & Freitas 1988, Pointier et al. 1991). In Lagoinha or Olhos d'agua lake - Seven species of mollusks were identified, each associated with one or more communities of macrophytes still present in this lake (Freitas et al. 1990). P. haustrum was the species that showed the highest density in the areas where the aquatic macrophytes had almost totally been destroyed, but it was present in the other communities too. In the degraded areas, its monthly average density was 41.6 +/- 15.1 individuals/m2 and 83% of its population were concentrated in habitats bearing no macrophytes. Analyses of the main biotic components in Lagoinha demonstrated that whereas P. haustrum was related positively (0.653) to the component "without macrophytes", B. straminea was related with the component "with macrophytes" (0.577). The density of B. straminea varied directly with the alteration in the biomass of Chara sp.(r = 0.849 and P< 0.05). In reference to this stand, B. straminea showed the highest monthly densities (55.5 +/- 41.9 individuals/m^2) occupying the second place in the communities of Eleodea sp. and Mayaca sp. from where the young individuals were collected in higher densities denoting a preferential area for reproduction of the species. The chemical characteristics of the waters of four sites of Lagoinha (Table I) are not very different from those of Lagoa Santa. Both of them present relatively low values of conductivity (ms 42,0 +/- 8,7 to 62,0 +/- 7,9), calcic hardness and alkalinity (mmol/l = 0,6 +/- 0,3 to 0,9 +/- 0,2), which are rather inferior to the ones found in springs and other lakes of the Lagoa Santa karstic system. INTERACTIONS BETWEEN B. TENAGOPHILA AND OTHER MOLLUSKS IN PAMPULHA RESERVOIR B. tenagophila and P. haustrum - In the Zoological area the densities of P. haustrum varied from about 40 to 480 specimens/m^2 in 1984/85. The increase in its density during March and May, however, did not evince a seasonal standard. In general, however, P. haustrum was present with two to ten times the population of the planorbids in this area during most of the period. It did not prevent the population growth of B. tenagophila, which came to surpass them for three times, during the last months of 1986 and 1987 (Fig. 2). In the Olhos d agua inlet, from 1986 to 1989 the densities of P. haustrum were similar to those found in the Zoo area, after its decline in 1987. B. tenagophila also underwent marked falls in 1987 and 1988, including sequences of negative months (zero) and months showing low densities, from 3.0 to 6.0 individuals/m^2. Data from monitoring on the borders of the reservoir evinced very uniform yearly averages of P. haustrum, varying monthly, however, from zero up to 34 snails/100 scoopfuls. The averages concerning B. tenagophila were lower (4.7 to 7.0) but the fluctuations were most pronounced with monthly peaks up to 62 snails. Table II shows the yearly averages of the snails all over the edge of the Pampulha reservoir. The data concerning P. haustrum at the Pampulha reservoir suggest that the eventual decline of B. tenagophila population, during a few months, can not result from competition with the pilid. But another hypothesis, that seems more consistent, is that density reductions of B. tenagophila and eventual disappearance of B. grablata, may be the result of conditions of habitat. These are often decisive factors in population dynamics (Jobim 1970, Pieri & Thomas 1987, Thomas 1987, Pointier et al. 1991). A third hypothesis would be the competitive action of M. tuberculata, recent invader, that has caused a fall of the density of P. haustrum and B. tenagophila throughout 1987. However, high densities of this thiarid in the following years, in other areas of the reservoir, did not hinder the coexistence and population increase either of P. haustrum and B. tenagophila which again bore high densities (Table II). B. tenagophila and B. grablata - Surveys carried out of the borders of the reservoir from 1953 to 1969 showed, in general, averages of B. glabrata per sample collection of less than 200 snails/year, in the whole reservoir. In the following eight years, the density of the B. glabrata population showed a sharp decline whereas B. tenagophila, recent invader, reached twice the density of B. glabrata. From 1981 to 1985 (Carvalho et al. 1985) B. glabrata was absent in the samplings for several months and the population was, on average, of about 428 B. tenagophila for each B. grablata or 18.000:1. Table III shows the average densities of the two planorbids. Population explosions in new reservoirs followed by sudden declines were observed mainly in the recently built reservoirs. This has been observed in about ten ponds and reservoirs by the authors (Freitas et al. 1992) and in almost a hundred ponds by Ruiz (1968), in Puerto Rico. These facts seem to be indicative of a lack of habitat conditions for B. glabrata in the reservoir (Freitas et al. 1987, Jordan et al. 1980). The success of biological control programmes in Puerto Rico may be partly due to the construction of small reservoirs, sometimes with destruction of the waterheads through silting, as we could observe in Itabira, MG. On the other hand, B. tenagophila seems to adapt well in reservoirs, considering they are the species' secondary habitat. In the Pampulha lake, the occurrence of B. tenagophila was stable. However, the density of the species varied, i.e., growing to the first great peak in 1981, but reaching the highest peak only in 1985. The growth pattern of B. tenagophila was quite different from that of B. glabrata and M. tuberculata. Whereas B. tenagophila reached higher densities only about ten years after having invaded a new reservoir, B. glabrata and M. tuberculata showed the highest peaks in the first years. The thiriad presented exponentional growth in each new reservoir invaded (Pampulha, Lagoa Santa, Lagoinha, Sumidouro, Rio Branco, etc.) as well as in each new site in these reservoirs, reaching in a short time about 10.000 up to 18.000 specimen/m^2. Is it possible that these qualities facilitate the competitivity among these mollusks? Some hypothesis have tried to explain the displacement of mollusks from their natural habitats. The introduction of B. glabrata in Manguinhos, Rio de Janeiro, resulted in the disappearance of B. tenagophila from this habitat. The disappearance of B. tenagophila was attributed to competitive exclusion by B. grablata (Magalh es 1966). Out of fifteen laboratory tests on coexistence between these two planorbidae, B. tenagophila was eliminated or strongly educed in twelve, and was partially successful in only one (Santos et al. 1989). B. grablata was displaced by B. tenagophila in the Horto Municipal and in the region of Hospital da Baleia, both located in Belo Horizonte, and was reduced strongly in the Pampulha reservoir (Paraense 1970, Carvalho et al. 1985). Its possible displacement by B. tenagophila in Ou- rinhos, SP (Kavazoe et al. 1980), was shown to be due to competitive exclusion. The authors concluded that B. glabrata had disappeared, however, the statistical analysis suggested a competition between the two species studied with a tendency to exclusion of B. glabrata. B. tenagophila and M. tuberculata - In the Zoological Garden area, B. tenagophila showed the usual population fluctuations: decline in the first half and peak in the last quarter of the year, independently of either the presence or the increase in density of P. haustrum since 1950 or M. tuberculata since 1986 (Tables I, II). The planorbids had shown very low densities (from 1.2 to 3.5 snails per m^2) for fifteen consecutive months, even before the invasion of M. tuberculata in this area, where it reached more than 800 thiarids/m^2. In the Olhos d'agua inlet, the density of the thiarid was incomparably higher, reaching a peak of over 16.000 snails/m^2 one year after the invasion. The fluctuation in density of B. tenagophila was rather similar in these two areas despite the different densities of the thiarid. In all the sites of the reservoir, searched at 1.0 and 2.0 m of depth, the densities of M. tuberculata remained high, from about 2.000 to 4.000 individuals/m^2, in the majority of the months (Fig.3). Nevertheless, in the Mergulh o mouth the densities of the thiarid were the highest in the reservoir, averaging 7.618 +/- 2.337 snails/m^2 (Bede 1992). B. tenagophila was concentrated in the mouth of the streams. Few years later (1988-93) due probably to the increase of pollution, the planorbid started to present the highest densities in the nearest areas to the barrage, an area less polluted. However, there was no correlation between M. tuberculata and several chemical factors in the water but there was negative correlation between the thiarid and rains (r = -0.682) in collections away from the border, at Mergulh o inlet. In the monitoring carried out all over the edge, the densities varied from 920 to 2.400 thiarids/100 scoopfuls. There was slight fluctuation and it could not be identified any marked seasonal factor concerning M. tuberculata. Nevertheless, B. tenagophila showed rather higher densities from August to October, before the rains. The ratio between M. tuberculata and B. tenagophila from 1987 to 1993 was about 200 to 400 thiarids for each single planorbid. On the edges, the percentage of dead thiarids (1989/91) was from about 20 to 30%, while in the transects perpendicular to the border, the death rates were from 50 to 90%. It should be emphasized that during the first year following the invasion (Carvalho 1986), the percentage of live thiarids was superior to 90% (Freitas et al. 1987). Different characteristics of the thiarids seem to enable it to be efficient competitors, such as extended life span, long- lasting high densities, capacity to compete for space and food and resistance to chemical factors released into the waters (Prentice 1983, Pointier & McCullough 1989, Bedˆ 1992). Promising results obtained by the introduction of T. granifera and M. tuberculata in several countries of Central America, have stimulated many scientists to suggest their use in biological control (Ferguson 1972, Malek & Malek 1978, Pointier et al. 1991). In their revision, Pointier and McCullough (1989), and Madsen (1990) presented many examples of the competitive ability of the thiarid. In Pampulha, however, eight species of mollusks, including B. tenagophila and P. haustrum, coexisted with M. tuberculata for about nine years without being apparently molested, except in the invasion year of each site of the dam. In the majority of the dams, lakes and ponds, when M. tuberculata arrived, there was neither B. grablata nor B. straminea as in Lagoa Santa, and the thiarid did not invade the streams. INTERACTIONS BETWEEN B.TENAGOPHILA AND B. GLABRATA IN STREAMS OF THE PAMPULHA BASIN The presence of B. grablata in the eight tributary streams at this natural area of its distribution, in the Pampulha basin, after its first collection by Martins and Versiani (1938), was confirmed by Martins and Falc o (1953) and by Paulini et al. (1967). But the presence of B. tenagophila or B. straminea coexisting with B. grablata in these streams, was detected two times at a ten year interval (Souza et al. 1981, Freitas et al.1991) (Table IV). While B. tenagophila was generally found at the six new sites, five of them near the mouth of the streams, B. grablata was rarely collected at the sites in the streams near the reservoir (Freitas et al. 1991). How to explain why B. tenagophila has not been able to establish itself in the streams although several attempts have been made for more than 20 years? (a) could it be due to B. glabrata that eliminate it from the streams? (b) or could the reason be the lack of rheotaxic capacity (Jurberg et al. 1988) which difficultates going upstream? (c) conditions of the habitat could be another hypothesis? We need more adequate parameters to define habitat conditions, collection stations, as well as to define coexistence. The presence of two species, for two or three years, in a biotope, may not signify coexistence but attempts of fixation or survival. B. glabrata continues to colonize all the brooks of the reservoir, swampy and marshy areas, principally in the streamheads which possibly are its breeding place or primitive habitat. When carried to the reservoir by rainfall, this species in general, disappears, but sometimes it can even colonize some areas, temporarily. B. straminea can be found up today in the same habitat and same site at Barauna stream, where it was collected by Souza et al. (1981) and is still coexisting with B. glabrata. These two species must have been carried to the reservoir by rainfall, although B. grablata was seldom collected at the reservoir and B. straminea has never been collected there or in the Barauna inlet. Among the unfavourable factors to these species could be the presence of fish in the reservoir, such as the Geophagus brasiliensis (Godinho et al. 1982), considered an efficient predator of planorbids (Weinztl & Jurberg 1990), or the presence of competitive mollusks, or even the elevated and growing organic pollution of the last twenty years. In the last case, the level of pollution might determine the minimum conditions of habitat for each species. A large area of the Ressaca/Sarandi streams in the mouth of the reservoir, with several months without OD (or = zero), can obstruct the passage of planorbids and fish, their predators. Besides, anaeroxy and elevated eutrophyzation levels (Giani et al. 1988, Pinto Coelho 1992) may be adverse conditions for planorbids (Branco & Matheus 1983, Freitas et al. 1992). However, the values of other chemical parameters did not seem to be a limiting factor to the snails (Baptista & Jurberg 1993). However, it can be supposed that if the conditions of habitat were favorable to B. straminea, it would try to grow in Barauna inlet when dragged by the flood and would at least bear some generations there. Such being the case, it would probably be collected there, at least occasionally in the reservoir (as B. glabrata was before 1986), from one of the thousands snail scoopfuls withdrawn per month, for several years. The coexistence among these planorbids is possibly more frequent in urban reservoirs, but examples of coexistence in natural environments are very rare. Paraense (1970) observed two cases of coexistence in Minas Gerais, and Teles (1989) reported the coexistence of B. glabrata and B. tenagophila only in 7 out of 1062 lots of planorbids collected in S o Paulo and coexistence of B. straminea and B. tenagophila in 12 lots. ACKNOWLEDGEMENTS To the scholarship holders Adriana Assis de Abreu, Adriano Mol Freitas, Giovana Fernandes Amorim, Marcos Antonio Reis Araujo, Simone Santos Mol and Valeria Rodrigues de Oliveira, for their participation in laboratory and field works. TABLES TABLE I Average density (ind/m^2) and maximum limits (p< 0.95) of four species of mollusks which were more abundant in four collection stations in the period of March 1987 to June 1988 in Lagoinha and Olhos d'agua Collection/ 1 2 3 4 stations Mollusks (Chara sp.) (without E. squamigera Mayaca and macrophytes) Elodea sp. -------------------------------------------------------------------------- Pomacea haustrum 9.5 +/- 5.9 41.6 +/- 15.1 3.4 +/- 4.5 0.2 +/- 0.4 Pisidum sp. 74.1 +/- 104.9 4.2 +/- 4.4 273.9 +/- 311.4 132.7+/- 186.8 Biomphalaria straminea 55.5 +/- 41.9 5.4 +/- 2.8 0 21.4+/- 21.3 Drepanotrema anatinum 0 0.3 +/- 0.4 31.2 +/- 24.1 23.0+/- 23.3 TABLE II Average of snails per 100 scoopfuls, per year, and minimum and maximum density^a in three years, on all the border of the Pampulha reservoir 1989 1990 1991 a P haustrum 9.9 10.3 8.3 (6.1 to 12.6) B. tenagophila 4.7 5.1 7.0 (2.1 to 21.3) M. tuberculata 1,420 1,780 1,490 (920 to 2.400) TABLE III Averages of Biomphalaria glabrata and Biomphalaria tenagophila per 100 scoopfuls, on the borders of the Pampulha reservoir, from 1981 to 1985 (The data until 1984 are based in Carvalho et al. 1985) Year 1981 1982 1983 1984 1985 Species (Aug/Sept) (Jan/Nov) (Jan/Nov) (Oct/Dec) (Jan/Oct) ----------------------------------------------------------------------- B. glabrata 0 0.04 3.20 0.012 0.025 B. tenagophila 4.50 2.30 17.90 10.31 10.27 TABLE IV Distribution of Biomphalaria species in the streams of the Pampulha basin, according to data by Souza et al.1981 and Freitas et al. 1991 Species Sites with the planorbids (Number) 1981 1991 --------------------------------------------------- B. grablata 18 12 (plus 6 new sites) B. strata inea 1a 1a B. tenagophila 1a 1a (plus 6 new sites)REFERENCES Andrade RM 1959. O problema da esquistossomose mansoni no lago artificial da Pampulha, Belo Horizonte, Minas Gerais (Brasil). Rev Bras Malar 10: 653-674. Andrade RM, Carvalho OS 1979. Colonizac o de Pomacea haustrum (Reeve, 1856) em localidade com esquistossomose mansoni: Baldim, MG, Brasil. (Prosobr. Pilidae) Rev Saude Publica S o Paulo 13: 92-107. Baptista DF, Jurberg P 1993. Factors conditioning the habitat and the density of Biomphalaria tenagophila (Orbigny, 1835) in an isolate schistosomiasis focus in Rio de Janeiro city. Mem Inst Oswaldo Cruz 88: 457-464. Barbosa FS 1973. Possible competitive displacement and evidence of hybridization between two Brasilian species of planorbid snails. Malacologia 14: 401-408. Bede LC 1992. Dinƒmica populacional de Melanoides tuberculata (Prosb.: Thiaridae) no Reservatorio da Pampulha, Belo Horizonte, MG, Brasil. MSc thesis. ICB - UFMG, Belo Horizonte. 112 pp. Branco SM, Mateus EM 1983. Desenvolvimento de caramujos planorbideos em lagoas de estabilizac o de esgostos. Rev DAE, S o Paulo 135: 37-42 Carvalho OS, Guimar es CT, Massara CL, Bonesio JER 1985. Situac o atual da esquistossomose mansoni no lago da Pampulha, Belo Horizonte - MG, Brasil. Rev Saude Publ SP 19: 270-278. Ferguson FF, Ruiz-Tiben E 1971. Review of biological control methods for schistosome-bearing snails. Ethiop Med J 9: 491-493. Ferguson FF 1972. Biological Control of Schistosomiasis Snail p. 85-92. In Schistosomiasis, Proceeding of a Symposium on the Future of Schistosomiasis Control. Tulane University, New Orleans, La, USA. Fradsen F, Madsen H 1979. A review of Helisoma duryi in biological control. Acta Tropica 36: 67-84. Freitas JR 1976. Ecologia de Vetores de Doencas, o Habitat Primitivo da Biomphalaria glabrata. Ci Cultura 28: 212-217. Freitas JR, Bede LC, De Marco Jr P, Rocha LA, Santos MBL 1987. Population Dynamics of Aquatic Snails in Pampulha Reservoir. Mem Inst Oswaldo Cruz 82: 299-305. Freitas JR, Alves CV, De Marco Jr P, Rocha LA, Santos MBL 1990. Preferˆncia de habitat de Biomphalaria straminea. Ci Cultura 42: 367-368. Freitas JR, Santos MBL, Lima LZ, Rocha LA, Souza CF 1991. Distribuic o dos moluscos hospedeiros intermediarios do Schistosoma mansoni na bacia da Pampulha, Belo Horizonte - atualizac o de dados. Ci Cultura 43: 641-43. Freitas JR, Santos MBL, Rocha LA 1992. Situac o atual da transmiss o da esquistossomose na Pampulha - Ecologia dos moluscos da represa. Anais do Seminario da Bacia Hidrografica da Pampulha: 41-70. Belo Horizonte, MG. Giani A, Pinto-Coelho RM, Oliveira SJM, Pelli A 1988. Ciclo sazonal de parametros fisicos quimicos da agua e distribuic o horizontal de nitrogˆnio e fosforo no reservatorio da Pampulha (Belo Horizonte, MG, Brasil). Ci Cultura 40: 69- 334. Godinho HP, Santos GB, Alves CBM, Formagio PS 1992. Os peixes e a pesca na Represa da Pampulha, Belo Horizonte, MG. Anais do Seminario da Bacia Hidrografica da Pampulha: 86-98. Belo Horizonte, MG. Guimar es CT 1978. Observac es bio-ecolologicas sobre Pomacea haustrum (Revee,1856). Sua utilizac o no controle biologico da esquistossomose mansoni. MSc thesis, Univ Fed Minas Gerais, Belo Horizonte - MG, 145 pp. Jobin WR 1970. Population dynamics of aquatic snails in three farm ponds of Puerto Rico. Am J Trop Med Hyg 19: 1038- 48. Jordan P, Christie JD, Unrau GO 1980. Schistosomiasis transmition with particular reference to possible ecological and biological methods of control. Acta Tropica 37: 95-135. Jurberg P, Coelho da Silva CLPA, Barreto MGM, Soares MS 1988. Rheotaxis of Biomphalaria glabrata on vertical substrates and its role in the recolonization of habitats treated with moluscicides. Mem Inst Oswaldo Cruz 83: 165-174. Kawazoe U, Magalh es LA, Hotta LK, Takaku L 1980. Competic o entre Biomphalaria glabrata (Say, 1818) e Biomphalaria tenagophila (d'Orbigny, 1835) em criadouros naturais no municipio de Ourinhos, SP (Brasil). Rev Saude Publ SP 14: 65-87. Madsen H 1990. Biological methods for the control of freshwater snails. Parasitol Today 6: 237- 241. Magalh es LA 1966. Estudo de uma populac o de Biomphalaria glabrata (Say, 1818) recentemente introduzida no Estado da Guanabara. Folha Clinica Biologica 35: 102- 120. Malek EA, Malek RR 1978. Potencial biological control of schistosomiasis intermediate hosts by helisome snails. The Nautilus 92: 15-18. McArthur RH 1972. Geographical Ecology. Harper and Row, New York, 269 pp. Martins AV, Versiani W 1938. Schistossomose mansoni em Belo Horizonte. Brasil Medico 52: 471-472. Martins AV, Falc o AL 1953. Distribuic o geografica dos planorbideos e seus indices de infestac o pelas cercarias do Schistosoma mansoni no municipio de Belo Horizonte. In Cong Bras de Higiene 10§, Curitiba. Anais. Nguma JS, McCullough FS, Masha E 1982. Elimination of Biomphalaria pfeifferi, Bulinus tropicus and Lymnea natalensis by the ampullarid snail, Marisa cornuarietis, in a man-made dam in northern Tanzania. Acta Tropica 30: 85-90. Paraense WL 1970. Planorbideos hospedeiros do Schistosoma mansoni. In AS Cunha, Esquistossomose Mansoni. Editora USP, S o Paulo. Paraense WL, Santos JM 1953. Um ano de observac o sobre esquistossomse em planorbideos da Lagoa Santa. Rev Bras Malariol D Tropicais 5: 243-269. Paulini E, Dias EP, Fiuza H 1967. Contribuic o a epidemiologia da esquistossomose em Belo Horizonte. Rev Bras Malariol D Tropicais 19: 571-606. Piere OS, Thomas JD 1987. Snail host control in the Eastern coastal areas of north-east (NE) Brazil. Mem Inst Oswaldo Cruz 82: 197-201. Pinto Coelho RM 1992. Evoluc o do grau de eutrofizac o na Lagoa da Pampulha: comparac o de ciclos sazonais de nutrientes (N e P) e organismos planct“nicos. Anais do Seminario da Bacia Hidrografica da Pampulha: 1-15. Belo Horizonte. Pointier JP, Theron A, Imbert-Establet, Borel 1991. Eradication of a sylvatic focus of Schistosoma mansoni using biological control by competitor snails. Biological Control 1: 244-247. Pointier JP, McCullough F 1989. Bilogical control of the snail hosts of Schistosoma mansoni in the Caribean area using Thiara spp. Acta Tropica 46: 147-155. Prentice MA 1983. Displacement of Biomphalaria glabrata by the snail Thiara granifera in field habitats in St. Lucia West Indies. Ann Trop Med Parasitol 77: 51-59. Ruiz E 1968. El control biologico de Biomphalaria glabrata con referencia especial a Marisa cornua- rietis. In Controle Ambiental da Esquistossomose Cap. VII:1-19. OPS, OMS. Publ Centro Eng Sanitaria UFMG. Belo Horizonte. Broch. 330pp. Ruiz-Tiben E, Palmer JR, Ferguson FF 1969. Biological control of Biomphalaria glabrata by Marisa cornuarietis in irrigation ponds of Puerto Rico. Bull Wld Hlth Org 41: 329-333. Santos MBL, Freitas JR 1988. Preference of Biomphalaria tenagophila (Gastrop.) among macrophytes and their periphytons determined through the degree of attractiveness. Rev Inst Med Trop S Paulo 29: 76-79. Santos MBL, Freitas JR, Alves ESR, Rocha LA 1989. Estrategia competitiva entre Biomphalaria glabrata e Biomphalaria tenagophila. Mem Inst Osw Cruz 84: 159- 179. Souza CP, Pereira JP, Rodrigues MS 1981. Atual distribuic o geografica dos moluscos hospedeiros intermediarios do Schistosoma mansoni em Belo Horizonte, MG, Brasil. Mem Inst Osw Cruz 76: 383-391. Sturrock RE 1974. Ecological notes on habitats of the freshwater snail Biomphalaria glabrata, intermediate host of Schistosoma mansoni. Caribean J Science 14: 149-162. Teles HM 1989. Distribuic o de Biomphalaria tenagophila e B. occidentalis no Estado de S. Paulo (Bra- sil). Rev Saude Publ SP S o Paulo, 23: 244-253. Thomas JD 1987. An evaluation of the interaction between freshwater pulmonate snail hosts of human schistosomiasis and macrophytes. Philosophical Tran R Soc London B 315: 75-125. Tilman D 1982. Resource Competition and Community Structure. Princeton, New Jersey. Princeton Uni. Press. 293pp. Weinztl M, Jurberg P 1990. Biological control of Biomphalaria tenagophila (Mollusca, Planorb.), a schistosomiasis vector, using the fish Geophagus brasiliensis (Pisces, Cichl.) in the laboratory or in a seminatural environment. Mem Inst Oswaldo Cruz 85: 35-38. Copyright 1995 Fundacao Oswaldo Cruz (Fiocruz)
The following images related to this document are available:Line drawing images[oc95052c.gif] [oc95052b.gif] [oc95052a.gif] |
| |||||||||

{kind=link}
{kind=link}