 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias Instituto Oswaldo Cruz, Vol. 90(2):281-284 mar./apr. 1995 A Molecular Genetic Study of the Variations in Metabolic Function During Schistosome Development Patrick J Skelly, Charles B Shoemaker Department of Tropical Public Health, Harvard School of Public Health, 665 Huntington Avenue, Boston, MA 02115, USA
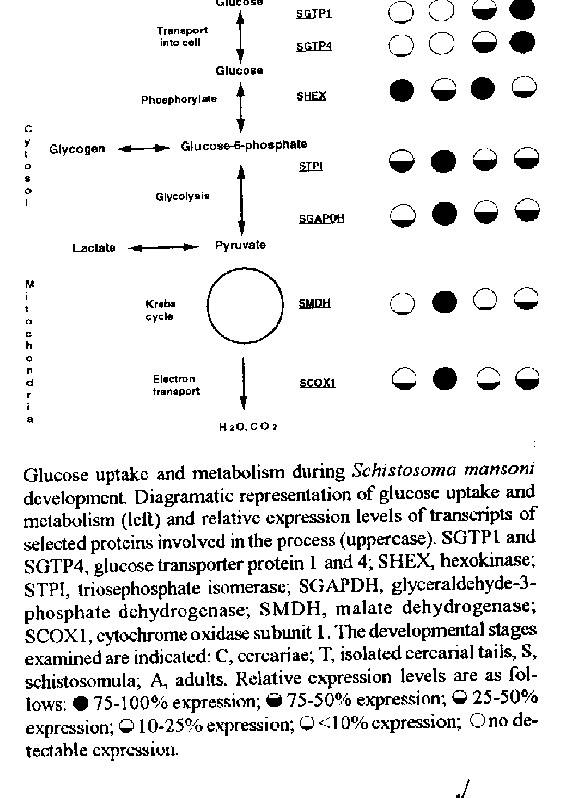
During their complex life cycle schistosomes alternate between the use of stored glycogen and reliance on host glucose to provide for their energy needs. In addition, there is dramatic variation between the relative contribution of aerobic versus anaerobic glucose metabolism during development. We have cloned a set of representative cDNAs that encode proteins involved in glucose uptake, glycolysis, Kreb's cycle and oxidative phosphorylation. The different cDNAs were used as probes to examine the expression of glucose metabolism genes during the schistosome life cycle. Steady state mRNA levels from whole cercariae, isolated cercarial tails, schistosomula and adult worms were analysed on Northern blots and dot blots which were quantified using storage phosphor technology. These studies reveal: (1) Transcripts encoding glycogen metabolic enzymes are expressed to much higher levels in cercarial tails than whole cercariae or schistosomula while the opposite pattern is found for glucose transporters and hexokinase transcripts; (2) Schistosomula contain low levels of transcripts encoding respiratory enzymes but regain the capacity for aerobic glucose metabolism as they mature to adulthood; (3) Male and female adults contain similar levels of the different transcripts involved in glucose metabolism. Key words: schistosome - metabolism - transcription - glucose During their development, schistosomes undergo profound morphological and biochemical change. Free-living cercariae emerge from their invertebrate snail host into fresh water where they employ oxidative glucose metabolism to provide their high energy needs as they are propelled by motile tails in their efforts to find and penetrate a final, vertebrate host (Van Oordt et al. 1989). During the invasion of this host, the cercariae lose their tails and transform into schistosomula. Biochemical studies suggest that this transformation is accompanied by a transition from an aerobic to a more anaerobic energy metabolism (Von Kruger et al. 1978, Thompson et al. 1984, Van Oordt et al. 1989). In the vertebrate blood stream the schistosomula develop into adults which generate large amounts of lactate (Schiller et al. 1975) but have been suggested to regain a greater reliance on aerobic metabolism (Coles 1972, Van Oordt et al. 1985). Schistosomes begin the process of glucose metabolism by transporting glucose across the tegument using a facilitated diffusion mechanism (Isseroff et al. 1972, Uglem & Read 1975, Cornford et al. 1988). Two full-length glucose transporter cDNAs (SGTP1 and SGTP4) have been obtained and shown to promote facilitated diffusion of glucose when expressed in Xenopus oocytes (Skelly et al. 1994). Functional characterization of these proteins in oocytes shows that the properties of SGTP1 and SGTP4 are similar to those reported for whole schistosomes (Skelly et al. 1994). Once inside cells, glucose is phosphorylated by hexokinase which prevents it from diffusing out of the cell. Hexokinase has been reported to be the rate-limiting enzyme in schistosome glucose metabolism (Shapiro & Talalay 1982, Tielens et al. 1985) and its cDNA has been cloned (Shoemaker et al. 1994). The phosphorylated glucose (glucose-6- phosphate) may be broken down by glycolysis through pyruvate to lactate or it may be stored as glycogen. The Schistosoma mansoni enzymes STPI and SGAPDH are involved in the glycolysis of glucose-6-phosphate and their cDNAs have been cloned (Shoemaker et al. 1992, Goudot-Crozel 1989). The complete breakdown of glucose to water and carbon dioxide occurs following the import of pyruvate into the mitochondria and catabolism via Krebs (TCA) cycle and electron transport. S. mansoni cDNA encoding enzyme representatives of the Krebs cycle (SMDH) and oxidative phosphorylation (SCOX1) are available (Skelly et al. 1993). Much of our understanding of the carbohydrate metabolism of schistosomes has been determined biochemically by examining the end products of absorbed, radiolabelled glucose. The present study uses the tools of molecular biology, together with the newly available cDNA probes, to examine the expression of the genes associated with glucose uptake and metabolism during the transition of S. mansoni cercariae into adults. MATERIALS AND METHODS Parasites - A Puerto Rican strain of S. mansoni was maintained by passage through Biomphalaria glabrata snails and CBA/J mice. RNA preparation and transcript analysis - RNA was recovered from whole cercariae, isolated cercarial tails, mechanically transformed schistosomula and adult male and female worms as previously described (Skelly et al. 1993). RNA from each stage was resolved by formamide gel electrophoresis and blotted to Nytran or dotted directly onto the membrane. Northern and RNA dot blots were hybridized with DNA probes (below) and the signal intensity was quantified using storage phosphor technology as previously described (Skelly et al. 1993). DNA isolation and probe production - Coding DNA fragments from genes encoding proteins involved in glucose uptake and metabolism were isolated as follows. In some cases, a fragment was amplified by the polymerase chain reaction (PCR) from adult or cercarial S. mansoni cDNA using degenerate oligonucleotides based on evolutionarily conserved amino acid sequence. This was done for the two glucose transporter proteins (SGTP1 and SGTP4) (Skelly et al. 1994), hexokinase (SHEX) (Shoemaker et al. 1994) and the mitochondrial proteins malate dehydrogenase (SMDH) and subunit 1 of cytochrome oxidase (SCOX1) (Skelly et al. 1993). A fragment of the glyceraldehyde-3-phosphate dehydrogenase (SGAPDH) gene was amplified by PCR from adult cDNA using oligonucleotides which were made based on the published sequence of this cDNA (Goudot-Crozel et al. 1989) and a fragment of the triosephosphate isomerase (STPI) gene was amplified from the complete, cloned STPI cDNA (Shoemaker et al. 1992). All PCR fragments were cloned and characterized by sequencing or restriction mapping to ensure their identity. The fragments were radiolabelled by the method of Feinberg and Vogelstein (1983) using sequence specific primers rather than random primers. RESULTS We have previously obtained DNA fragments encoding proteins involved in glucose uptake (SGTP1 and SGTP4, Skelly et al. 1994), phosphorylation (SHEX, Shoemaker et al. 1994), glycolysis (STPI, Shoemaker et al. 1992, SGAPDH, Goudot-Crozel et al. 1989) and respiration (SMDH, SCOX1, Skelly et al. 1993). Each of these cDNAs has been utilized as a probe with which to examine transcript levels encoding these proteins in cercariae, schistosomula and adults. In most cases the relative amounts of each transcript was assessed by quantitative RNA dot blots which were performed several times and analyzed statistically (Skelly et al. 1993). For the glucose transporters, Northern blots were scanned using a densitometer to estimate relative transcript abundance (Skelly et al. 1994). The results of these studies on glucose metabolism gene expression at different developmental stages is depicted diagramatically in the Figure, right panel. Filled circles identify stages containing the highest level of transcript detected in our studies; less-filled circles depict lower message levels and empty circles indicate no detectable signal. Both glucose transporter mRNAs are poorly expressed in whole cercariae (and are undetectable in cercarial tails) but are readily detectable in schistosomula and at their highest level in adults. SHEX mRNA levels are very high in both whole cercariae and schistosomula and much lower in cercarial tails and adults. Transcripts encoding the enzymes of glycolysis and oxidative metabolism show the highest levels of expression in cercarial tails with low to moderate levels of expression in whole cercariae, schistosomula and adults. Importantly the levels of expression of the mitochondrial enzymes are relatively higher in adults compared with schistosomula (or whole cercariae). DISCUSSION In this report we summarize our knowledge of developmental changes in the carbohydrate metabolism of S. mansoni as assessed by changes in gene expression. We have cloned part or all of several cDNAs encoding proteins pivitol in the uptake and utilization of glucose by these parasites and used them to assess steady state mRNA levels during the cercaria to adult transformation. While transcript levels clearly represent the capacity to express their encoded proteins, it is important to note that regulation may also occur during translation and through protein turnover and, thus, mRNA abundance may not reflect the amounts of the encoded proteins. Even variations in the abundance of metabolic gene products cannot always be interpreted in terms of their affect on glucose metabolism since some proteins, such as GAPDH, have been shown to perform separate functions unrelated to metabolism (Singh & Green 1993). In spite of these potential reservations, changes in transcript levels reported in this study do largely corroborate biochemical investigations of glucose metabolism in schistosomes and help resolve some controversy regarding adult metabolism. Cercariae - Cercariae exhibit very low levels of glucose transporter mRNAs and this is presumably a reflection of the fact that these organisms in fresh water will likely not encounter exogenous glucose. As expected, hexokinase is poorly expressed in cercarial tails since cercariae derive energy from glycogen breakdown which does not require this enzyme. The transcript profile of isolated cercarial tails is consistent with a prominant aerobic metabolism there - high levels of SMDH and SCOX1 transcripts as well as of the glycolytic enzymes examined (Bruce et al. 1971, Von Kruger et al. 1978, Van Oordt et al. 1989). This supports the view that the primary function of the tail is to generate energy for propulsion. The overall transcript profile of whole cercariae is similar to that of schistosomula (with the notable exception of the glucose transporter proteins). This suggests that cercarial bodies, like schistosomula and unlike cercarial tails, engage in anaerobic glucose catabolism to a large degree. Why cercariae should express such a high level of SHEX, though, is a puzzle in view of the low SGTP transcript levels. Perhaps the SHEX mRNAs are stored in cercariae in preparation for immediate use upon penetration of the final host and the transformation to schistosomula begins. Schistosomula - One of the primary roles of the invading schistosomula must be the uptake of host glucose to supplant depleted reserves of glycogen and to provide energy for subsequent development. Commensurate with this hypotheseis is the increased expression of SGTP genes in schistosomula to fulfill this function. At this stage, high SHEX levels are important for phosphorylating the newly acquired glucose to prevent it from diffusing back out and to prepare it for catabolism. Schistosomula which take up glucose in vitro produce copious quantities of lactate indicating a dominant anaerobic metabolism at this development stage (Thompson et al. 1984, Van Oordt et al. 1989). High levels of glycolytic transcripts and low levels of SMDH and SCOX1 transcripts observed in schistosomula is consistant with this view. Adults - In agreement with the fact that adults consume prodigious quantities of glucose, the highest levels of the two SGTP messages are seen at this stage. The SHEX level, in contrast, is surprisingly low. One explanation could be that the SHEX mRNA detected here is a larval hexokinase and that a second, adult-specific hexokinase exists to phosphorylate the large amounts of glucose taken in by the mature parasites. However, the comparable hexokinase kinetic measurements obtained using either larval or adult extracts, as well as the partial purification and N-terminal sequencing of an adult hexokinase and its identity to the deduced N-terminal sequence of SHEX, strongly suggests that the larval and adult enzymes are the same (Tielens et al. 1994). Perhaps a more likely ex- plaination is that the low level of SHEX mRNA in adults reflects the reported role of hexokinase as the rate-limiting glycolytic enzyme in schistosomes (Shapiro & Talalay 1982, Tielens et al. 1985). In this study, it is clear that high SHEX transcript levels correlate with lower aerobic metabolism whereas those developmental stages with low SHEX transcript levels exhibit a higher degree of aerobic metabolism. It may be that SHEX plays a central role in controling the cells response to absorbed glucose and in the establishment of this pattern. While the glycolytic transcript amounts in schistosomula and adults are comparable, the levels of the mitochondrial transcripts are higher in adults. This is consistant with more recent data showing that a significant proportion of adult energy is derived through respiration, which requires mitochondrial function and results in the generation of carbon dioxide and water (Van Oordt et al. 1985). The experiments with adults were conducted on separated males and females. Because the transcript levels of both were very similar the results are presented together. Despite the fact that females produce enormous quantities of eggs, a process which requires a good deal of energy, our findings suggest that the metabolic enzyme transcription profiles of both males and females are similar. REFERENCES Bruce JI, Ruff MD, Hasegawa H 1971. Schistosoma mansoni: Endogenous and exogenous glucose and respiration of cercariae. Exp Parasit 29: 86-93. Coles GC 1972. Oxidative phosphorylation in adult Schistosoma mansoni. Nature 240: 488-489. Cornford E M, Fitzpatrick AM, Quirk TL, Diep CP, Landaw EM 1988. Tegumental glucose permeability in male and female Schistosoma mansoni. J Parasitol 74: 116-128. Feinberg AP, Vogelstein B 1983. A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal Biochem 132: 6-13. Goudot-Crozel V, Caillol D, Dhabali M, Dessein AJ 1989. The major parasite surface antigen associated with human resistance to schistosomiasis is a 37-kD glyceraldehyde-3P- dehydrogenase. J Exp Med 170: 2065-2080. Isseroff H, Bonta CY, Levy MG 1972. Monosaccharide absorption by Schistosoma mansoni - I. Kinetic characteristics. Comp Biochem Physiol 43A: 849-858. Schiller EL, Bueding E, Turner VM, Fisher J 1975. Aerobic and anaerobic carbohydrate metabolism and egg production of Schistosoma mansoni in vitro. J Parasitol 61: 385-389. Shapiro TA, Talalay P 1982. Schistosoma mansoni: Mechanisms in regulation of glycolysis. Exp Parasitol 54: 379-390. Shoemaker CB, Gross A, Gebremichael A, Harn D 1992. cDNA cloning and functional expression of the Schistosoma mansoni protective antigen, triose phosphate isomerase. Proc Natl Acad Sci USA 89: 1842-1846. Shoemaker CB, Reynolds SR, Wei G, Tielens AGM, Harn DA 1994. Schistosoma mansoni hexokinase: cDNA cloning and immunogenicity studies. Exp Parasitol 80: 36-45. Singh R, Green MR 1993. Sequence-specific binding of transfer RNA by glyceraldehyde-3-phosphate dehydrogenase. Science 259: 365-368. Skelly PJ, Stein LD, Shoemaker CB 1993. Expression of Schistosoma mansoni genes involved in anaerobic and oxidative glucose metabolism during the cercariae to adult transition. Mol Biochem Parasitol 60: 93-104. Skelly PJ, Kim JW, Cunningham J, Shoemaker CB 1994. Cloning, characterization and functional expression of cDNAs encoding glucose transporter proteins from the human parasite, Schistosoma mansoni. J Biol Chem 269: 4247- 4253. Tielens AGM, Houweling M, van den Bergh SG 1985. The effect of 5-thioglucose on the energy metabolism of Schistosoma mansoni in vitro. Biochem Pharmacol 34: 3369- 3373. Tielens AGM, van den Heuvel JM, van Mazijk HJ, Wilson JE, Shoemaker CB 1994. The 50-kDa, glucose-6-phosphate sensitive hexokinase of Schistosoma manson: Enzymatic properties and amino-acid sequence analysis. J Biol Chem 269: 24736-24741. Thompson DP, Morrison DD, Pax RA, Bennett JL 1984. Changes in glucose metabolisma nd cyanide sensitivity in Schistosoma mansoni during development. Mol Biochem Parasitol 13: 39-51. Van Oordt BEP, van den Heuvel JM, Tielens AGM, van den Bergh SG 1985. The energy production of the adult Schistosoma mansoni is for a large part aerobic. Mol Biochem Parasitol 16: 117-126. Van Oordt BE, Tielens AGM, van den Bergh SG 1989. Aerobic to anaerobic transition in the carbohydrate metabolism of Schistosoma mansoni cercariae during transformation in vitro. Parasitology 98: 409-415. Von Kruger WMA, Gazzinelli G, Figueiredo EA, Pellegrino J 1978. Oxygen uptake and lactate production by Schistosoma mansoni cercaria, cercarial body and tail, and schistosomule. Comp Biochem Physiol 60B: 41-46. Uglem GL, Read CP 1975. Sugar transport and metabolism in Schistosoma mansoni. J Parasitol 61: 390-397. Copyright 1995 Fundacao Oswaldo Cruz (Fiocruz) The following images related to this document are available:Line drawing images[oc95055a.gif] |
| |||||||||

{kind=link}