 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias Instituto Oswaldo Cruz, Vol. 90(2):311-318 mar./apr. 1995 Calomys callosus: an Alternative Model to Study Fibrosis in Schistosomiasis Mansoni. The Pathology of the Acute Phase JA Lenzi+, EM Mota, M Pelajo-Machado, RAN Paiva, HL Lenzi Departamento de Patologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
[TABLES AT END OF TEXT]
Twenty Calomys callosus, Rengger, 1830 (Rodentia -
Cricetidae) were studied in the early stage of the acute
schistosomal mansoni infection (42nd day). The same number of
Swiss Webster mice were used as a comparative standard. Liver
and intestinal sections, fixed in formalin-Millonig and
embedded in paraffin, were stained with hematoxilin and eosin,
PAS-Alcian Blue, pH=1,0 and 2,5, Lennert's Giemsa, Picrosirius
plus polarization microscopy, Periodic acid methanamine
silver, Gomori's silver reticulin and resorcin-fuchsin.
Immunohistological study (indirect immunofluorescence and
peroxidase labeled extravidin-biotin methods) was done with
antibodies specific to pro-collagen III, fibronectin, elastin,
condroitin-sulfate, tenascin, alpha smooth muscle actin,
vimentin and desmin. The hepatic granulomas were small, reaching only 27% of the volume of the hepatic Swiss Webster granuloma. They were composed mainly by large immature macrophages, often filled by schistosomal pigment, characterizing an exsudative-macrophage granuloma type. The granulomas were situated in the parenchyma and in the portal space. They were often intravascular, poor of extracellular matrix components, except fibronectin and presented, sometimes alpha smooth muscle actin and vimentin positive cells. The C. callosus intestinal granulomas were similar to Swiss Webster, showing predominance of macrophages. Therefore, the C. callosus acquire very well the Schistosoma mansoni infection, without developing strong hepatic acute granulomatous reaction, suggesting lack of histopathological signs of hypersensitivity. Key words: Calomys callosus - Schistosoma mansoni - fibrosis - liver - intestine - granuloma - extracellular matrix Schistosomiasis research lacks an experimental model, particularly in small laboratory animals, that reproduces with accuracy the human disease, justifying the search of new ones. Calomys callosus, Rengger, 1830 (Rodentia - Cricetidae), a wild mouse-like autochthonous rodent from South America (Argentina, Bolivia, Paraguay), is found also in Brazil in the Northeast, Central-west and Southern Regions (Moojen 1952, Hershkovitz 1962, Massoia & Fornes 1965, Mello 1969, Mello & Moojen 1979). Petter et al. (1967) introduced C. callosus as a new laboratory rodent. Further biological studies have shown that this animal is easy to handle and to adapt to the laboratory conditions, and has high fertility rates during the whole year (Justines & Johnson 1970, Mello 1977, 1978). Johnson et al. (1965) and Justines and Johnson (1969) have experimentally infected C. callosus with Machupo virus, thus establishing its importance as an experimental model. Borda (1972) and Mello (1979, 1980) have shown experimentally that C. callosus is a permissible host for Schistosoma mansoni and Coelho et al. (1979) found 7.70 % C. expulsus (same as C. callosus according to Hershkovitz 1962) to be naturally infected in the region of Caratinga, State of Minas Gerais, Brazil. In this paper we are presenting the major histopathological changes that occur in the liver and intestines of C. callosus during the early stage of the acute schistosomal infection, using Swiss Webster mice as a comparative standard. MATERIALS AND METHODS Twenty C. callosus and twenty Swiss Webster mice were infected when they were five days old by percutaneous exposure to 70 cercariae of the Belo Horizonte isolate of S. mansoni, and were sacrificed on the 42nd day of infection. Tissue samples were taken from the liver and intestines, fixed in formalin-Millonig (Carson et al. 1973), and embedded in paraffin. Intestinal samples were prepared according to Swiss roll procedure (Lenzi & Lenzi 1986). Sections were stained with hematoxilin and eosin; PAS-Alcian Blue, pH = 1,0 and 2,5, Lennert's Giemsa; Picrosirius plus polarization microscopy (Junqueira et al. 1979), Periodic Acid Methanamine Silver (PAMS), Gomori's silver reticulin and resorcin-fuchsin. The size of hepatic granulomas was determined in histological section of six animals per group, stained with hematoxilin and eosin. The diameters of ten granulomas containing single egg in the center were measured in each liver using an ocular micrometer. Granuloma volume was calculated assuming a spherical shape (Cheever & Barral-Netto 1985). To study specific intra and extracellular matrix components, cryostat or trypsinized liver sections were studied by indirect immunofluorescence or indirect peroxidase labeled extravidin-biotin staining (ExtrAvidin-Peroxidase, Sigma E- 2886; Anti-mouse polyvalent immunoglobulins biotin conjugate (Sigma, B-2016); Anti-rabbit immunoglobulins biotin conjugate (Sigma, B-3275). Anti-tenascin and anti-mouse IgG, IgA and IgM were tested with immunofluorescence and the other antisera with immunoenzimatic method: Pro-Collagen III (Institut Pasteur, Lyon); Fibronectin (Dakopatts, A-082); Condroitin- sulfate (Sigma, C-8035); Elastin (Sigma, E-4013); Vimentin (Sigma, V-7505); Desmin (Sigma, D-9284) and alpha smooth muscle actin (Sigma, A-2547). Normal sera were used as control, plus secondary antibody-FITC or secondary antibody- biotinylated-extravidin-peroxidase. Diaminobenzidine (DAB) was used as chromogen (DAB: 3mg; TBS: 10 mL; 30% H2O2: 10 uL). Sections were coverslipped in buffered glycerin with p- phenylenediamine (Johnson et al. 1982) and examined with a Zeiss microscope equipped with epifluorescence system. RESULTS
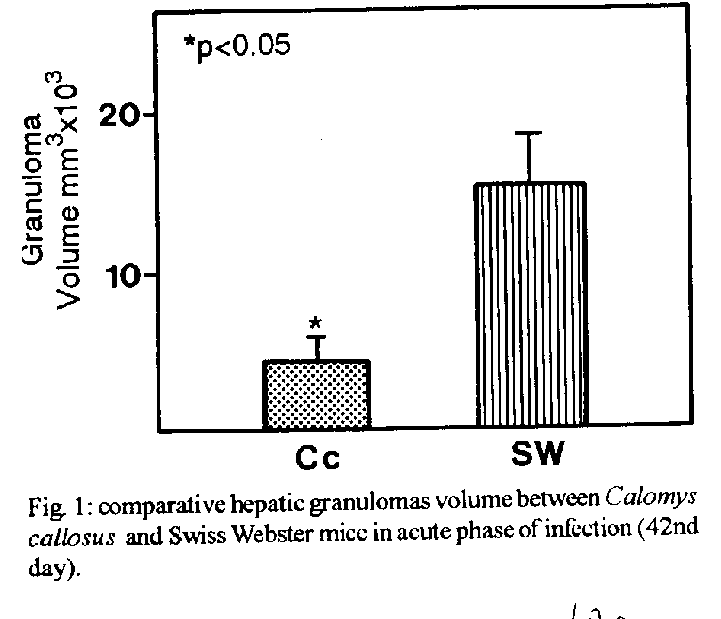
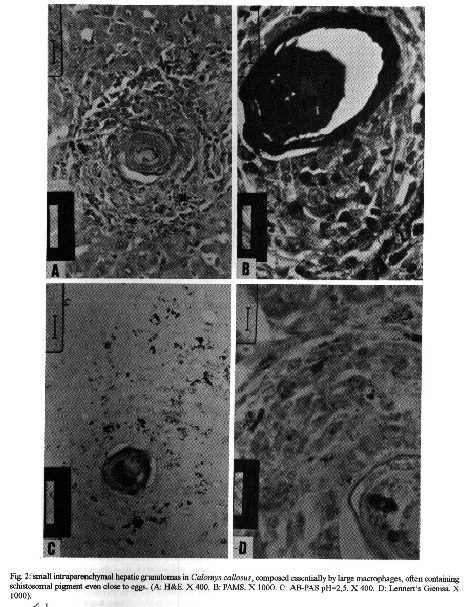
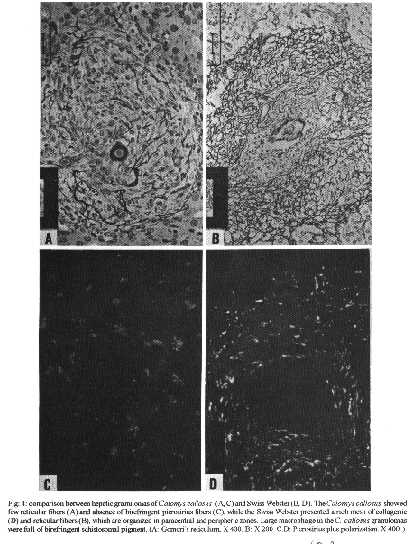
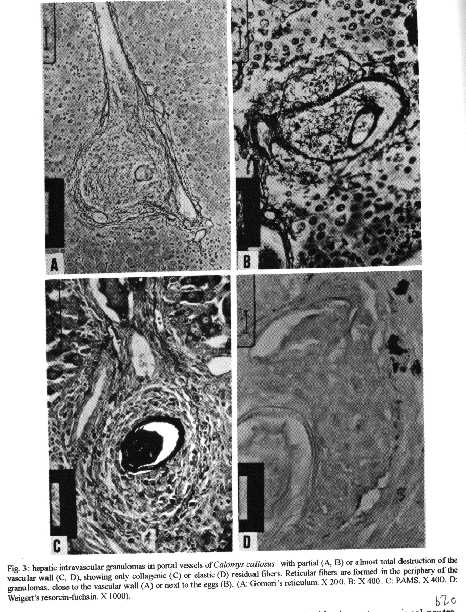
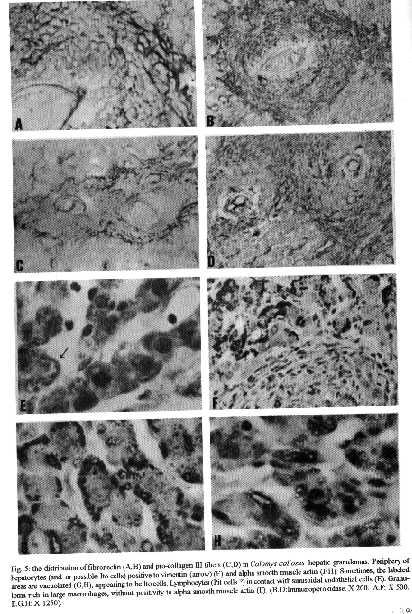
The hepatic granulomas of C. callosus were small, reaching only 27 % of the volume of hepatic Swiss Webster granulomas, used as controls (Fig. 1). They were predominantly composed by large and mature macrophages, often filled by schistosomal pigment (Fig. 2), and less number of eosinophils and neutrophils, and scarce lymphocytes. Mast cells were detected only in advanced S.Webster exsudative-productive granulomas. Unlike Swiss Webster hepatic granulomas, they were less zonally configured, and presented very few reticular and birefringent picrosirius fibers (Fig. 4). The smaller granulomas were situated in the parenchyma and the larger ones in the portal space. These were often intravascular, externally demarcated by residual vascular wall elastic and reticular fibers (Fig. 3). The granuloma extracellular matrix components were better developed by immunohistology, showing the presence, in both group of animals, of fibronectin and pro-collagen III, which in the C. callosus were usually localized in the periphery of the granulomas (Fig. 5 A-D). The C. callosus granulomas never exhibit an external zone with myeloid metaplasia as was seen in Swiss Webster hepatic ones. Periovular diffusion of PAS positive material was more common in Swiss Webster than C. callosus granulomas. Focal parenchymal necrosis was found in both group of animals, and portal inflammation and fibrosis was absent or minimal in the C. callosus liver. These presented marked Kupffer cell hypertrophy, with cells enlarged and filled with granular schistosomal pigment. While the Swiss Webster hepatocytes were alpha smooth muscle actin and weakly vimentin positive, the C. callosus hepatocytes showed higher amount of alpha-actin and intermediate filaments in the following order of intensity: alpha smooth muscle actin > vimentin > desmin (Fig. 5 E-H). It was impossible to discriminate Ito cells by these antisera due to their striking labeling in the periphery of the hepatic cells. Some portal granulomas in both group of animals exhibited cells positive to alpha smooth muscle actin and vimentin. The antibodies against mouse immunoglobulins did not react with C. callosus immunoglobulins. The tenascin and condroitin-sulfate were negative in both group of animals and elastin was detected only in vessel walls. In C. callosus intestines, there was large number of eggs within the mucosa and submucosa vessels. The intestinal granulomas, predominantly located in the mucosa layer, were also rich in macrophages, with less number of eosinophils and neutrophils. In the mucosal lamina propria there was a diffuse infiltration of eosinophils and some mast cells not related to the granulomas. The main hepatic and intestinal histopathological findings are schematized in Tables I-III. DISCUSSION The Swiss Webster hepatic granulomas, at this time of infection (42nd day) showed the classical aspects described by Hsu et al. (1972), with predominance of initial exsudative- productive type. The C. callosus hepatic granulomas, on the contrary, presented peculiar aspects characterized by prevailing intravascular location, development of scarce or null fibrosis, and predominance of matures and large macrophages, many of them containing schistosomal pigment, characterizing a peculiar exsudative-macrophage granuloma type. This small granuloma pattern is unlike the mouse down- regulated granulomas, which exhibit more fibrosis, suggesting different mechanisms involved in their functions. The small parenchymal granulomas are similar to BALB/C nude mice granulomas as was described by Byram and Lichtenberg (1977). They observed that the hepatic lesions in the nude mice were much smaller and the lacked epithelioid macrophage, with lesions about mature eggs typically consisting of monocytes and macrophages filled with pigment, occasional neutrophils, and rarely one or more eosinophils or giant cells. However, the C. callosus appear to be efficient in circumscribing the egg antigens, because there was very limited periovular diffusion of PAS positive material. The C. callosus hepatic lesions were also more or less like to those described in Nectomys squamipes by Rodrigues-e-Silva (1989), which is also a cricetidae. It is important to stress the absence of perigranulomatous myeloid metaplasia concomitant with absence of an external zone, which was usually present in Swiss Webster hepatic granulomas, presenting a peripheric mesh of reticular fibers, rich in pro-collagen III and fibronectin, that functions as a medullary stroma (Fig. 4-B). The presence of alpha smooth muscle actin and vimentin positive cells in some portal granulomas of both group of animals, suggests the participation of myofibroblasts in the granuloma composition. The C. callosus intestinal lesions, except the diffuse eosinophil infiltration in the mucosal lamina propria and predominance of macrophages in the granulomas, was similar to those observed in mice. The frequent presence of intravascular granulomas in the C. callosus liver may be due to a more limited periovular inflammatory reaction than in Swiss Webster. We suggested that the passage of the eggs from the vessels to the tissues, as the passage of the eggs to the intestinal lumen, is dependent on inflammatory cells surrounding the eggs (Lenzi et al. 1991). Indeed, Raso et al. (1983) observed in the liver of thymectomised and S. mansoni infected mice a marked inhibition of the development of granulomas, scarce inflammatory exsudation around the parasite eggs and many intravascular eggs. It appears that C. callosus acquire very well the schistosomal infection, as was demonstrated by Borda (1972), Mello (1979) and Coelho et al. (1979) without developing strong acute granulomatous response. Therefore, we detected an excellent model to study low acute responsiveness to egg antigens in the liver, which forms granulomas that, in contrast to Swiss Webster, lack histopathological signs of hypersensivity, without being an athymic animal.
TABLE I
Liver: comparative histopathologic lesions between
Calomys callosus and Swiss Webster mice infected
with Schistosoma mansoni (42 days of infection)
Findings C callosus Swiss Webster
Findings C. callosus Swiss Webster
-------------------------------------------------------------
Granuloma:
- stage "E" (macrophage E - P
- size Small Large
Granuloma composition:
- Macrophages ++ ++++
- Epithelioid cells +/- -
- Giant cells - -
- Neutrophils +/- +/-
- Eosinophils +/++ ++++
- Fibroblastoid cells -/+ +
- Mast cells - -/+
- Lymphocytes +/- +/-
- Plasmocytes - -
E = Exsudative; E-P = Exsudative-Productive; (+ to
++++) = Levels of intensity;
(-) = Negative; (+/-) = Weakly positive; (-/+) = Nega-
tive or positive.
TABLE II
Liver: comparative histopathologic lesions between
Calontys callosus and Swiss Webster mice infected
with Schistosoma tnansoni (42 days of infection)
Findings C. callosus S. Webster
--------------------------------------------------------------
Granuloma ECM:
- Reticulin Fibers +/- +/+++
- Collagen Type I -/+ +
- Collagen Type III + ++
- Fibronectin ++ ++
- Tenascin - -
- Elastin - -
- Condroitin-sulfate _
- Alpha-smooth muscle
Actin -/+ -/+
- Vimentin -/+ -/+
- Desmin - -
- Pigmented Macrophages +++ +/-
Extramedullary metaplasia - ++
Extragranulomatous changes:
Portal inflammation +/- ++
Portal fibrosis - +
Kupffer hyportrophy with
pigment +++ +
Focal necrosis + +
Adult worms ++ +/-
ECM = Extracellular Matrix; (+ to +++) = Levels of intensity;
(-) - Negative; (+/-) - Weakly positive; (-/+) Negative or
positive.
TABLE III
Intestine: comparative histopathologic lesions
between Calomys callosus and Swiss Webster mice
infected with Schistosoma mansoni (42 days of
infection)
Findings C.callosus Swiss Webster
-------------------------------------------------------
Granuloma:
- stage Exsudative Exsudative
- size Small Large
- location Mucosa Mucosa
Granuloma composition:
- Macrophages ++ ++
- Eosinophils ++ +++
- Neutrophils +
Mucosa:
- Eosinophil infil-
tration Diffuse Localized
- Mast cells +/- -
(+ to +++) = Levels of intensity, (-) = Negative; (+/-) =
Weakly positive; (-/+) = Negative or positive.
ACKNOWLEDGMENTS To AS Rodrigues, FF Cruz, H Ferreira, ID Pedro, LFG Caputo and VC Valentin for technical assistance and to HMN Diniz, GJ Vieira, JC Cruz and VCR Sa for preparing the figures. REFERENCES Borda CE 1972. Infecc o natural e experimental de alguns roedores pelo Schistosoma mansoni Sambon, 1907. Master thesis, UFMG, Belo Horizonte, 43pp. Byram JE, Lichtenberg FV 1977. Altered schistosome granuloma formation in nude mice. Am Trop Med Hyg 26: 944-956. Carson FL, James MS, Martin JH, Lynn JA 1973. Formalin fixation for electron microscopy: A re-evaluation. Am J Clin Path 59: 365-373. Cheever AW, Barral-Netto M 1985. Fibroblast stimulating activity of extracts of hepatic granulomata of Schistosoma mansoni-infected rodents with marked or slight hepatic fibrosis. Trans R Soc Trop Med Hyg 79: 319-321. Coelho PMZ, Dias M, Mayrink W, Magalh es P, Mello MN, Costa CA 1979. Wild reservoirs of Schistosoma mansoni from Caratinga, an endemic schistosomiasis area of Minas Gerais state, Brazil. Am J Trop Med Hyg 28: 163-164. Hershkovitz P 1962. Evolution of neotropical cricetine rodents (Muridae) with special reference to Phyllotine group. Fieldiana Zool 46: 1-524. Hsu SYL, Hsu HF, Davis JR, Lust GL 1972. Comparative studies on the lesions caused by eggs of Schistosoma japonicum and Schistosoma mansoni in livers of albino mice and Rhesus monkeys. Ann Trop Med Parasitol 66: 89-97. Johnson GD, Davidson RS, McNamee KC, Russel G, Godwin D, Holbrow EJ 1982. Fading of immunofluorescence during microscopy: a study of the phenomenon and its remedy. J Immunol Meth 55: 231-242. Johnson KM, Mackenzie RB, Webb PA, Kuns ML 1965. Chronic infection of rodents by Machupo virus. Science 150: 1618-1619. Junqueira LCU, Bignolas G, Brentani RR 1979. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem J 11: 447-455. Justines G, Johnson KM 1969. Immune tolerance in Calomys callosus infected with Machupo virus. Nature 222: 1090-1091. Justines G, Johnson KM 1970. Observations on the laboratory breeding of the cricetine rodent Calomys callosus. Lab Anim Care 20: 57-60. Lenzi HL, Lenzi JA 1986. Swiss-roll technique examination of intestines in experimental animals. Rev Soc Bras Med Trop 19: 106. Lenzi HL, Lenzi JA, Kerr IB, Antunes SLG, Mota EM, Oliveira DN 1991. Extracellular matrix in parasitic and infectious diseases. Mem Inst Oswaldo Cruz 86: 77-90. Massoia E, Fornes A 1965. Nuevos dados sobre la morfologia, distribucion geografica y etoecologia de Calomys callosus callosus (Rengger) (Rodentia - Cricetidae). Physis (Buenos Aires) 25: 325-331. Mello DA 1969. Roedores silvestres de alguns municipios do estado de Pernambuco e suas regi es naturais. Rev Bras Pesq Med Biol 2: 360-362. Mello DA 1977. Observac es preliminares sobre a ecologia de algumas especies de roedores do cerrado, municipio de Formosa, Goias, Brasil. Rev Bras Pesq Med Biol 10: 39-44. Mello DA 1978. Biology of Calomys callosus (Rengger, 1830) under laboratory conditions (Rodentia, Cricetinae). Rev Bras Biol 38: 807-811. Mello DA 1979. Infecc o experimental de Calomys callosus (Rengger, 1830), (Cricetidae - Rodentia) a quatro especies de parasitos. Rev Soc Bras Med Trop 13: 101- 105. Mello DA 1980. Estudo populacional de algumas especies de roedores do cerrado (norte do municipio de Formosa, Goias). Rev Brasil Biol 40: 843-860. Mello DA, Moojen J 1979. Nota sobre uma colec o de roedores e marsupiais superiores de algumas regi es do cerrado do Brasil central. Rev Bras Pesq Med Biol 12: 287-291. Moojen J 1952. Os roedores do Brasil. Instituto Nacional do Livro. Biblioteca Cientifica Brasileira, Serie A- II, 123pp. Petter F, Karimi Y, Almeida CR 1967. Un nouveau rongeur de laboratoire, le cricetide Calomys callosus. C R Acad Sc Paris 265: 1974-1976. Raso P, Rocha OA, Pereira LH, Tafuri WL 1983. Efeito da timectomia neonatal na esquistossomose mansoni experimental. Rev Soc Bras Med Trop 16: 112-121. Rodrigues-e-Silva R 1989. Nectomys squamipes e Akodon arviculoides (Rodentia:Cricetidae) como hospedeiros naturais do Schistosoma mansoni em Sumidouro (RJ- Brasil). Emprego do Nectomys como modelo alternativo no estudo da esquistossomose mansoni. Master thesis - Instituto Oswaldo Cruz - FIOCRUZ, Rio de Janeiro, 147pp. Copyright 1995 Fundacao Oswaldo Cruz (Fiocruz)
The following images related to this document are available:Halftone images[oc95062e.gif] [oc95062c.gif] [oc95062d.gif] [oc95062b.gif]Photo images[oc95062b.jpg] [oc95062d.jpg] [oc95062c.jpg] [oc95062e.jpg]Line drawing images[oc95062a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}