 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Genetic Variability and Differentiation between Populations of Rhodnius prolixus and R. pallescens, Vectors of Chagas' Disease in Colombia German Lopez, Jaime Moreno* Unidad de Bioquimica, Centro Intemacional de Entrenamiento e Investigaciones Medicas (CIDElM), A.A. 5930, Cali, Colombia Departamento de Biologia, Universidad de Antioquia, A.A. 1226, Medeillin, Colombia
Enzyme polymorphism in Rhodnius prolixus and R. pallescens (Hemiptera, Reduviidae), principal vectors of Chagas' disease in Colombia, was analyzed using starch gel electrophoresis. Three geographic locations were sampled in order to determine gene flow between populations and to characterize intra- and interspecific differences. Of 25 enzymes assayed 10 were successfully resolved and then used to score the genetic variation. The enzymes PEPD, GPI, PGM and ICD were useful to differentiate these species and PGD, PGM and MDH distinguished between sylvatic and domiciliary populations of R. prolixus. Both polymorphism and heterozygosity indicated greater genetic variability in sylvatic habitats (H= 0.021) compared to domiciliary habitats (H= 0.006) in both species. Geneflow between sylvatic and domiciliary populations in R. prolixus was found to be minimal. This fact and the genetic distance between them suggest a process of genetic isolation in the domiciliary population. Key words: Rhodnius prolixus - Rhodnius pallescens - isoenzyme polymorphism - Chagas' disease - Triatominae - Colombia The blood-sucking bug Rhodnius prolixus Stal (Hemiptera, Reduviidae) is the' main vector of Trypanosoma cruzi, the causative agent of Chagas' disease, in Colombia, Venezuela and Central America (Lent & Wygodzinsky 1979). This disease is a public health problem throughout Latin America, where it has been estimated that 16-18 million people are infected (WHO 1990). One of the main problems in the control of T. cruzi vectors is the reeolonization of domiciliary habitats through the migration of triatomine bugs between palm trees and human dwellings (Gomez-Nunez 1969, Dujardin et al. 1991). To detennine the mobility between sylvatie and domiciliary habitats it is informative 10 analyze the genetic structure of representative populations with respect to gene flow. Enzyme systems provide useful genetic markers for population studies, allowing the genetic structure to be elucidated on the basis of polymorphic loci. Isoenzyme polymorphism has been used to characterize populations of Triatoma infestans, the principal vector of T. cruzi in Bolivia. One study suggested a founder effect during the expansion of the populations (Dujardin & Tibayrenc 1985). In addition, no significant differences in enzyme phenotype were observed be- tween sylvatic and domiciliary populations (Dujardin el al. 1987). In Chile, Frias and Kattan (1989) reported that the domiciliary T. infestarts show less enzymatic polymorphism than the sylvatic T. spinolai Porter. A similar approach was used to compare some species of the genus Rhodnius in Venezuela, where R. prolixus and R. robustus Larrouse were found to have identical isozyme patterns in spite of having evident morphological differences (Harry et al. 1992). The present study reports the genetic variability of 10 isoenzymes in sylvatic, domiciliary and peridomiciliary populations of R. prolixus and R. pallescens Barber. In addition, we analyze the rate of gene flow between sylvatic and domiciliary habitats. MATERIALS AND METHODS Insects - Three geographic locations were selected for the study (Fig. 1). The presence of triatomines in distinct habitats provided appropriate conditions to study gene flow and rendered feasible the characterization of intra- and interspecific genetic variability. Forty-one domiciliary and 43 sylvatic R. prolixus were collected around the vil- lage of Coyaima, department of Tolima (populations A and B, respectively); 36 sylvatic R. pallescens were collected around the Galeras, department of Sucre (population C) and 41 peridomiciliary R. pallescens in San Carlos, Antioquia (population D). Males and females were identified according to the key of Lent and Wygodzinsky (1979). COLOMBIA
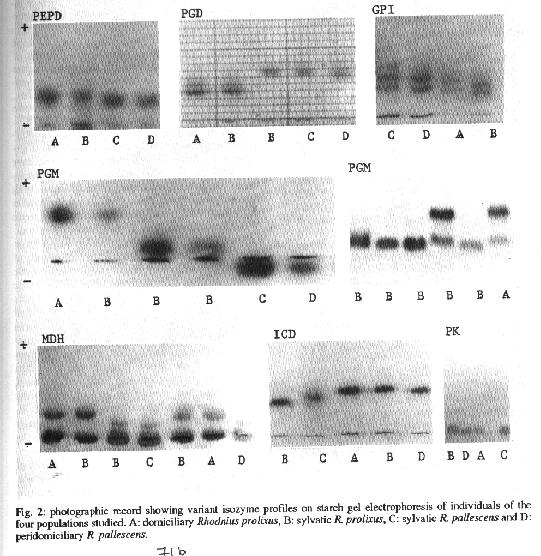
Electrophoresis and enzyme detection Standard horizontal starch gel electrophoresis procedures and enzyme development conditions described by Harris and Hopkinson (1976) and Miles et al. (1980) were employed, with the ex- ception of sample loading (10 ul/well), voltage reduction to avoid overhearing of gels and extension of running time to achieve resolution. The following 10 enzymes were assayed: aminopeptidase D (PEPD) E.C. 3.4.13.9; phosphogluconate dehydrogenase (PGD) E.C. 1.1.1.44; glucose phosphate isomerase (GPI) E.C. 5.3.1.9; phosphoglucomutase (PGM) E.C. 2.7.5.1; malate dehydrogenase (MDH) E.C. 1.1.1.37; isocitrate dehydrogenase (ICD) E.C. 1.1.1.42; pyruvate kinase (PK) E.C. 2.7.1.40; esterase (ES) E.C. 3.1.1.1; alanine aminotransferase (ALAT) E.C. 2.6.1.2 and aspartate aminotransferase (ASAT) E.C. 2.6.1.1. The electrophoretic patterns were recorded photographically. Data analysis - Genetic variability was calculated as the average of polymorphic loci (P) and mean heterozygosity (H). Gene flow was estimated by Fst, which is a measure of variation in allele frequencies among different populations. Specifically, Fst is the variance in allele frequency (Vq) standardized by the mean (q) (Fst -Vq/(q(t-q)) (Futuyma 1986). Hierarchical cluster analysis by the complete linkage method was utilized to determine similarty between populations (Duun & Everitt 1982). The coefficient of genetic distance between populations was defined as D=(l-cosO)1/2 and cosO- E(piA.piB)112 where cosO is a measusure of genetic distance between the two populations A and B, piA and piB being the gene frequencies for each allele at a given locus in the two populations (CavalliSforza & Edwards 1967). RESULTS The electrophoretic patterns of the isoenzymes PEPD, GPI, PGM and ICD distinguished between R. prolixus and R. pallescens and PGD, PGM and MDH between sylvatic and domiciliary populations of R. prolixus (Fig. 2). Phenotypes of PGD, MDH and PK were shared by sylvatic R. prolixus and R. pallescens. The enzymes ES, ALAT and ASAT showed monomorphic patterns for all the samples assayed in both species. However, the enzymatic activity in the latter was too weak or bands too diffuse to allow reliable interpretation. The polymorphic isozyme profiles are shown in Fig. 2. The sylvatic population of R. prolixus presented one phenotype for PEPD and GPI; two for PGD, MDH, ICD and PK and three phenotypes for PGM. In contrast, the domiciliary popu- lation of this same species presented only one polymorphic enzyme, PGM, having two phenotypes. Evidence for heterozygosity was found among sylvatic R; prolixus for the enzymes PGD, PGM and PK. Only PK was polymorphic among the sylvatic R pallescens analyzed, while peridomiciliary individuals of this species displayed monomorphic phenotypes for all the enzymes examined. We found no evidence of any correlation between enzymatic pattems and sex. The variability in the populations assayed was H=0.011 and P=9.09 for domiciliary R. prolixus; H=0.035 and P=72.7 for sylvatic R. prolixus; H=0.011 and P=18.2 for sylvatic R. oallescens.
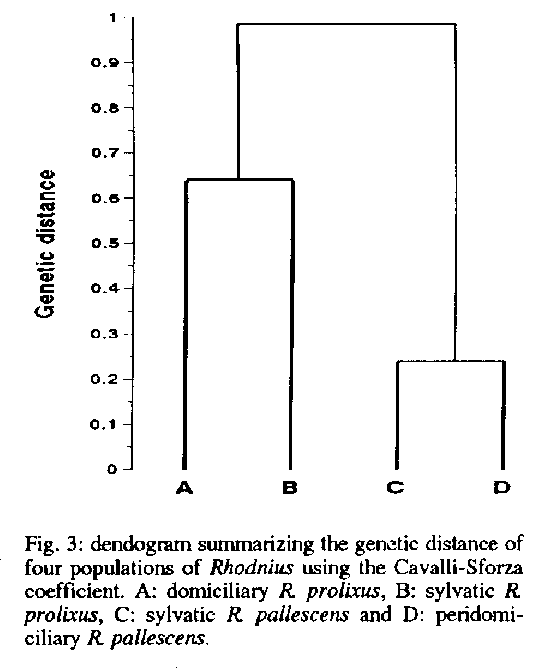
The gene flow between sylvatic and domiciliary populations of R. prolixus was minimal with FsT scores above 0.025 for five polymorphic loci (Table). The genetic distance between R. pallescens populations using the Cavalli-Sforza coeffi- cient was 0.236. A greater genetic distance of 0.636 was observed between sylvatic and domiciliary populations of R. prolixus. The genetic be 0.982. The genetic distance relationships are summarized in Fig. 3. TABLE Gene flow analysis between sylvatic and domiciliary populariotas of Rhodnius prolixus calculated with five polymorphic loci using genetic Wright's variance
---------------------------------------------- Locus^a FsT^b ---------------------------------------------- PGD 0.684 PGM 0.492 MDH 0.826 ICD 0.654 PK 0.323 ---------------------------------------------- Mean 0.598 ---------------------------------------------- a: only include loci that shown phenotypic differences between both populations b: genetic Wright's variance
The enzymes PEPD, PGD, GPI, PGM, MDH, ICD and PK had the most readily detectable activity and therefore were most suitable for genetic analysis of triatomine vectors. The PGM locus was routinely scorable for differentiation between R. prolixus populations in addition to comparisons between Rhodnius species. On the other hand, the enzymes PEPD, PGD, GPI, MDH and ICD appeared to be more useful in differentiating between species. The occurrence of three bands in phenotypes of GPI indicates a dimeric structure. This is consistent with the known quaternary structure for this enzyme (Harry et al. 1992). Our results indicate that the total genetic variability was low for the populations of both species analyzed at H=0.019 for R prolixus and H=0.005 for R. pallescens. Heterozygosity values of approximately 0.1 are usually recorded for invertebrates (Nevo 1983, Ayala and Kiger 1984). Nevertheless, in the present study the heterozygosity was almost four times greater in R. prolixus than R. pallescens. This is comparable with the observed genetic variability reported by Harp et al. (1992) for K prolixus from Venezuela where the heterozygosity ranged between 0.04 to 0.10. T. infestans from Bolivia showed heterozygosity values of 0.04 (Dujardin & Tibayrenc 1985). Such low genetic variability appears to be a common feature of specialist insects (Nevo 1983). Comparison of the variability between sylvatic and domiciliary or peridomiciliary vectors indicates greater variability in sylvatic than domiciliary populations. Hence the polymorphism in natural populations of R. prolixus could be used to indicate local and geographical variations of this species. The variety of biological niches available to vectors in sylvatic habitats may influence the maintenance of genetic variability, similar to the heterogeneity occurring in sylvatic stocks of T. cruzi studied by Saravia et al. (1987). The remarkable monomorphism in the domiciliary populations of R. prolixus suggests a genetic drift process. If this is true, the domiciliary populations of vectors could have been established by a small number of individuals with minimal gene flow. This possibility is consistent with the results of the analysis by genetic Wright's variance. The high values of FST indicate genetic isolation of domiciliary R. prolixus. Consequently, is not surprising that the strains of T. cruzi of domiciliary origin show characteristic genetic profiles quite different to those of sylvatic origin. This hypothesis is supported by results from Widmer et al. (1985) who showed that domiciliary and peridomiciliary T. cruzi stocks from geographically dispersed loci were pheno- typically uniform. In addition, enzyme polymorphism in 54 stocks of T. cruzi from vectors, mammalian reservoirs and infected humans, showed that the variability in loci of sylvatic transmission was greater than in loci of domiciliary transmission and the patterns of heterogeneity correlated with the type of transmission cycle, domiciliary or sylvatic (Saravia et al. 1987). Our findings are in accord with the results of radioisotope labelling of bugs in Venezuela, where migration between palms and houses was not evident (Gomez-Nunez 1969). On the other hand, the average rate of gene flow among established populations of a species is often quite low (Futuyma 1986). This is especially likely in triatomine bugs because the immigrating individuals must compete with residents to survive and reproduce. In addition, it appears that the sylvatic and domiciliary loci of R. prolixus were stable. Blood meal availability may be a determinant of the observed genetic isolation of these colombian populations. These findings suggest a founder effect in the domiciliary vectors and provide diagnostic loci for species and populations of the same species of Rhodnius. In practical terms the results support the feasibility of the prevention of recolonization using materials and construction designs that discourage the establishment of sylvatic populations in the domiciliary habitat. ACKNOWLEDGEMENTS To Dr Nancy Gore Saravia of the Fundacidn Centre Internacional de Entrenamiento e Invetigaciones Medicas (CIDElM), who provided helpful suggestions in the development of these studies. To Mrs Berta Velez for technical asistance. To Dr Juan Carlos Ortiz, Unidad de Bioquimica (CIDElM) and Dr Bruce Alexander, Unidad de Entomologia (CIDElM) for czitical wading of early drafts of this mannscript. This research was developed and presented in paflial fulfillment of the requirements for the Master of Science degree, Departamento de Biologia, Universidad de Antioquia, Medellin, Colombia. REFERENCES Ayala FJ, Kiger Jr JA 1984. Genetica moderna, Omega, Barcelona, Espana. p. 146-148. Cavalli-Sforza LL, Edwards AWF 1967. Phylogenetic analysis: models and estimation procedures. Evolution 21: 550- 570. Dujardin JP, Tibayrenc M 1985. Etude de 11 enzymes et donnes de genetique fonnalle pour 19 loci enzymatiques chez Triatoma infestans frlemimptera: Reduviidae) en Bolivia. J Med Trop 65:271-280.
Dujardin JP, Tibayrenc M, Venegas E, Maldonado L, Desjeux P, Ayala FJ 1987. Isozyme evidence of lack of speciation between wild and domestic Triatoma infestans (Hemiptera: Reduviidae) in Bolivia. J Med Ent 24: 40-45. Dujardin JP, Garcia-Zapata MT, Jurberg J, Roelants P, Cardozo L, Panzera F, Dias JCP, Sohofield CJ 1991. Which species of Rhodnius is invading houses in Brazil? Trans R Soc Trop Med Hyg 85: 679-680. Dunn G, Everitt BS 1982. An introduction to mathetnatical taxonomy. 2nd ed. Cambridge University Press, New York, U.S.A.p. 25-137. Frias D, Kattan F 1989. F. studio de taxonom/a molecular en poblaciones de Triatonza infestans (Klug, 1934) y Triatoma spinolai Poller 1933, (Hemiptera: Reduviidae). Act Ent Chil 15:205-210.
Futuyma DJ 1986. Evolutionary biology. 2nd ed. Sun- derland, Massachusetts, U.S.A.p. 129-148. Gomez-Nunez JC 1969. Resting places, dispersal and survival of Co^60-tagged adult Rhodnius prolixus. J Med Entomol 6: 83-86. Hams H, Hopkinson DA 1976. Handbook of enzyme electrophoresis in human genetics. 2nd ed. London, England, p. 1.1-3.16. Harry M, Galindez I, Cariou ML 1992. Isozyme variability and differentiation between Rhodnius prolixus, R. robustus and R pictipes, vector of Chagas' disease in Venezuela. Med Vet Ent 6: 37-43. Lent H, Wygodzinsky P 1979. Revision of Triatominae (Hemiptera: Reduviidae), and their significance as vectors of Chagas' disease. Bull Amer Mus Nat Hist NY163: 123- 520. Miles MA, Lanham M, De Sousa AA, Povoa M 1980. Further enzymic characters of Trypanosoma cruzi and their evaluation for strain identification. Trans R Soc Trop Med Hyg 74:231- 237.
Nevo E 1983. Population genetics and ecology: the interface, p. 231-287. In Evolution from molecules to men, Bendall, Cambridge U.K. Saravia NG, Holguin AF, Cibulskis RE, D'Alessandro A 1987. Divergent isozyme profiles of sylvatic and domiciliary Trypanosoma cruzi in the Eastem Plains, Piedinont and Highlands of Colombia. Am J Trop Med Hyg 36: 59-69. WHO 1990. Tropical Diseases 1990. TDRJCTD/ HH20.1 WHO Geneva, 26 pp. Widmer G, Marinkelle CJ, Guhl F, Miles MA 1985. Isozyme profiles of Trypanosotna cruzi stocks from Colombia and Ecuador. Ann Trop Med Parasit 79: 253-257.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95071b.gif]Photo images[oc95071b.jpg]Line drawing images[oc95071c.gif] [oc95071a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}