 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Repair of Schistosomal Intestinal Vascular Lesions after Curative Treatment Luciana Silva+, Zilton A Andrade Laboratorio de Esquistossomose, Centro de Pesquisas Gonctalo Moniz-FIOCRUZ, Rua Valdemar Falcao 121, 40295-001 Salvador, BA, Brasil
Code Number: OC95078
Size of Files:
Text: 17K
Graphics: Photos (jpg) 127K / Halftones (gif) 292K
The process of repairing intestinal vascular lesions
induced by schistosomiasis in mice was studied before and
after curative chemotherapy, by means of histopathology
coupled with injections of the mesenteric veins with India ink
or plastic, in this case followed by corrosion in strong
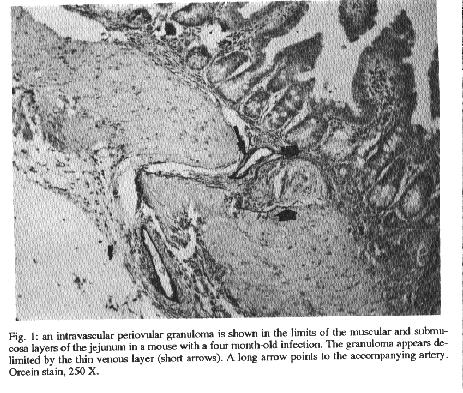
acid.The granulomas were avascular, mainly formed while within blood vessels, and were associated with distortion of the intestinal vasculature in their proximity, represented by tortuosities, focal dilatation, narrowing, and anastomosis of the mucosal and submucosal veins. Two to four months after cure of schistosontiasis involuting granulomas were seen to be slowly vascularized, a process going from the periphery toward the center of the granulomas. No intravascular granulomas were seen four months after treatment. The previously distorted mucosal and submucosal veins gradually regained their normal appearance, only a slight tortuosity remaining. Key words: Schistosoma mansoni - vascular lesions - repair - intestinal schistosomiasis The natural habitat of Schistosoma mansoni is the mesenteric veins of man and of several other susceptible mammals. The female worms preferentially lay their eggs within the terminal venous radicles of the intestines. Thus, to be excreted into the stools the schistosome eggs need first to mature and to pass through the vascular wall and interstitial tissues of the intestines where many of them are trapped and induce a granulomatons response. Since the infection with S. mansoni can last for long periods, a considerable number of eggs and accompanying granulomas can accumulated in the intestines and damage their vasculature. It has been demonstrated that more eggs are accumulated in the intestines than in the liver during S. mansoni infections of several hosts (Cheever 1969). Although clinical manifestations related to intestinal lesions in man are usually mild (Andrade & Silveira 1990), severe vascular lesions have been described in experimental schistosomal infections. These latter consisted of arte- riovenous shunts, tortuosity and dilatation of intestinal veins (Nagy et al. 1981) and obstruction and reversion of blood flow diverting eggs toward the serosal surface (Kloetzel 1971). Present study aims at investigating about these vascular lesions and how they could be modified by curative treatment. The way vascular lesions are repaired may be an important factor to be considered for the understanding of the peculiar behavior of intestinal schistosomiasis. MATERIALS AND METHODS One hundred outbred albino Swiss mice of both sexes, 20/23g, were infected transcutaneously, on the same day, with approximately 50 recently shed S. mansoni cercariae each. Two months after exposure all animals showed viable schistosome eggs in the stools. Half of the infected mice were killed two or four months after infection. The other half, then with a four-month old infection, was submitted to treatment with a combination of two drugs administered simultaneonsly by gavage: oxamniquine (100mg/kg bw) and praziquantel (100mg/kg bw). The treated animals were sacrificed one, two, three and four months after treatment. In addition, for the study of intestinal vasculature, one group of normal, non-infected mice was included. At the time of sacrifice the mice were anesthetized with ether and exsanguinated by severing of the abdominal aorta. For vascular studies the animals were kept alive up to the beginning of the portal vein injections. The following methods were applied: Parasitology - Cercariae were obtained from laboratory raised and infected Biomphalaria glabrata. Fecal examination for confirmation of infection was directly performed by the Kato's method. Since the intestines were utilized in totum for other methods, intensity of infection was evaluated only in the liver by the counting of eggs according to Cheever (1970). Histology - The intestines were open along the mesenteric border, emptied of its contents, turned into a roll and then fixed in 10% buffered formalin. The roll was kept in position during all the procedures for paraffin embedding and cutting, so all parts of the intestines were represented in a single microscopic section. These sections were stained with hematoxylin and eosin and, in especial cases, with PAS-Schiff, Picrosirius-red for collagen, and orcein for elastic fibers. Vascular injections (1) - Shortly before sacrifice, the mice were intraperitoneally injected with 200uu of heparin. A canula was introduced into the portal vein and the intestinal vessels were first perfused with a 5% solution of sodium cit- rate until clearing of the vasculature. Through the same canula the intestinal vessels were slowly filled under low pressure with India ink diluted 3:1 in 2.5% gelatin until a good and uniform filling of the intestinal vessels was obtained. After injection the intestines were removed on block, washed in tap water and opened along the mesenteric border. Porttions of the intestines were distended and fastened over a cardboard and left for fixation in 10% formalin. After a few days, selected paris of the intestines were dehydrated in alcohol and cleared in xylol. These flattened and semi-transparent portions of the intestine, pre- served in glycerol, were smashed between two slides and analyzed grossly and microscopically. As for the treated animals this procedure was only made four months after treatment. (2) Vinylite (vinyl acetate) in 7% acetone was used for vascular injection. The plastic solution was similarly injected into the portal vein and the animal kept in the refrigerator overnight. Then, the intestines were removed and placed in 50% hydrochloric acid. About one week later the plastic cast was taken out of the acid, washed several times in water and alcohol, dried, and examined directly and under a dissecting microscope. RESULTS All exposed animals became infected. Intensity of infection varied from mild to moderate. An average of 10,971 eggs per gram of liver tissue were found after four months of infection. In histological sections schistosomal periovular granulomas were observed along the entire length of the intestines, isolated or clustered, predominating in the ileal and colonic sub-mucosa. Also, there were granulomas formed in the mucosal and muscular layers, but only rarely in the sub-serosa. Granulomas were discrete, cellular, with scanty fibrous stroma, having macrophages as the main component, although some of them exhibited in addition many eosinophils. Elastic staining revealed that most of the granulomas were delimited by thin fibers of fragmented elastic tissue, indicating their intravascular location (Fig. 1). Distribution of granulomas was segrnental, and the areas devoid of them presented normal histology. By simple inspection of the sec- tions no clear-cut differences were observed in animals with two or four months of infection, regarding size, preferential location, cellular composition and quantity of intestinal periovular granulomas. Vascular injection with India ink revealed that the granulomas appeared as clear, non-injected, avascular areas. A fine network of anastomosing veins and capillaries was present at the periphery of some granulomas (Fig. 2). The straight, non-branching sub-mucosal vessels observed in the normal mouse, appeared tortuous, numerous, and with many collaterals in the vicinity of granulomas. Besides, these vessels pre- sented local diIatation alternating with local narrowing, twisting and short-circuits. The latter refers to dilated vessels making a loop and returning to near the point of origin. The characteristic vascular arrangement of the intestinal villi was usually preserved, even in the neighborhood of confluent granulomas. No arteriovenous shunts were detected in any of the preparations examined. Many animals had their mesenteric veins injected, but preparations of good quality were obtained in 5, 6, 7 and 15 cases of infected (two months), infected (four months), infected and treated, and control animals, respectively. Plastic cast of the intestinal vasculature were made in 25 animals, but yielded poor preparations, because the finest veins were not filled. The technique was not used for treated animals. Only the increased vascuIarity and tortuosity of submucosal veins were documented in infected animals. Three and four months after treatment intestinal granulomas became two to three times smaller than before and their total number diminished considerably as seen in histological sections. Granulomas were represented by a small collection of macrophages and a few eosinophils surrounding empty egg shells. None appeared intravascularly. They were seen in the interstitial tissue, near dilated small veins which presented an irregular outline, sometimes with minute points of focal destruction of their elastic layer (Fig. 3). ColIagen fibers appeared coarse and fragmented at the periphery of the shrunken granulomas. Within the muscular layer some hypertrophic smooth muscle fibers were seen near involuting collections of granulomas. The intestinal vasculature as seen in injection preparations in the treated mice retumed to.near normal in a span of four months. Shrunken granulomas remained avascular, but the peripheral capillary network became more and more
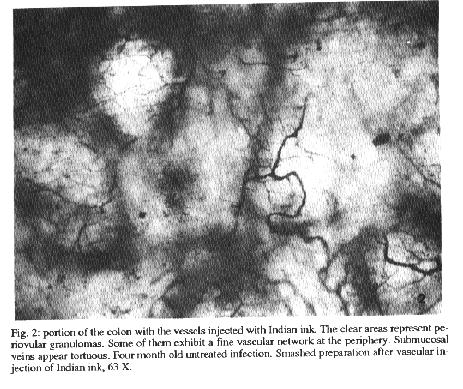
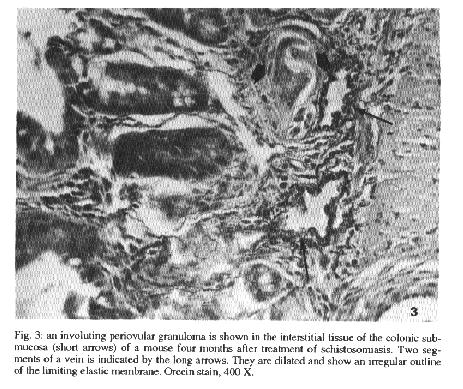
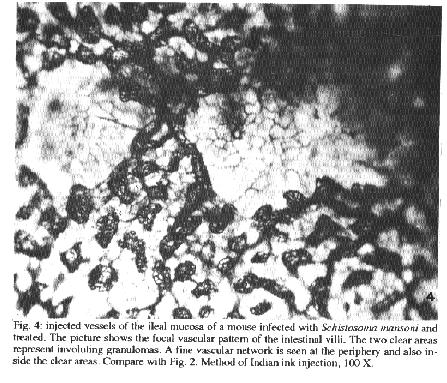
Fig. 2: potion of the colon with the vessels injected with Indian ink. The clear areas represent periovular granulomas. Some of them exhibit a fine vascular network at the periphery. Submucosal veins appear tortuous. Four month old untreated infection. Smashed preparation after vascular injection of Indian ink, 63X. Fig. 3: an involuting periovular granuloma is shown in the interstitial tissue of the colonic submucosa (short arrows) of a mouse four months after treatment of schistosomiasis. Two segments of a vein is indicated by the long arrows. They are dilated and show an irregular outline of the limiting elastic membrane. Orcein stain, 400 X. Fig. 4: injected vessels of the ileal mucosa of a mouse infected with Schistosoma mansoni and treated. The picture shows the focal vascular pattern of the intestinal villi. The two clear areas represent involuting granulomas. A fine vascular network is seen at the periphery and also inside the clear areas. Compare with Fig. 2. Method of Indian ink injection, 100 X. vascular threads were noted (Fig. 4). As for the usually straight mucosal and sub-mucosal veins only a slight degree of tortuosity persisted. Vascularization of the intestinal villi remained undisturbed DISCUSSION The mature eggs of S. mansoni seem to be less pathogenic to the intestines than to the liver or lungs. Although a considerable number of eggs may accumulate in the intestines during a prolonged infection, proportionally little clinical or pathological manifestations result therefrom. In a study of necropsy material including 502 S. mansoni infected Brazilian subjects and 933 noninfected, no statistically significant differences were found on the frequency of colitis, entero-colitis, polyps or intestinal fibrosis between the two groups (Cheever & Andrade 1967). Presumably, lesions are continuously being produced during egg elimination through the intestinal wall, especially involving its vasculature (Kloetzel 1971, Nagy et al. 1981). Peculiarities of the vasculature apart, some data point to a singular reactivity of the intestines: the periovular granulomas are immunologically modulated from the start (Weinestock & Boros 1981), being smaller than those in the liver and poorly fibrogenic (Santos et al. 1992). Data obtained from present investigation confirm that marked vascular changes occur in intestinal schistosomiasis of the mouse during recent infection. In addition, this study reveals that vascular lesions are rapidly repaired after cure. Considering these aspects changes are qualitatively similar to those in the liver (Andrade & Brito 1981), especially regarding new vascularization of the granulomas after cure. However, the present study failed to confirm two important changes previously described in the literature: the formation of arteriovenous shunts (Nagy et al. 1981) and the reversion of blood flow revealed by the shunting of eggs toward the serosal surface of the intestines (Klcetzel 1971). In the first case, changes apparently similar to those illustrated by Nagy et al. (1981) have been observed in our injection preparations, but were not interpreted as arteriovenons shunts. Probably, for the better evaluation of arterio-venous shunts it is necessary that arterial vessels be simultaneously injected, as did Nagy at al. (1981). In the second case, predominant distribution of eggs in the subserosal areas may be related to extremely heavy infection, as has been seen before (Andrade et al. 1974). In the present case, infection of moderate intensity might not have been sufficient to cause inversion of blood flow. Many intestinal granulomas were found intravasculafly in non-treated animals, two and four months after infection. After treatment, involuting granulomas were no longer observed within blood vessels, but in paravascular locations. Then, the veins and venules in close proximity to the granulomas usually appeared dilated and with focal alterations of their elastic membrane. It can be assumed that the vessel re-acquired its continuity, by-passing the lesions. Probably these changes occur spontaneously during the involution of the periovular granulomas. Curative treatment only accelerates these changes and make them synchronous throughout. Of course there is much more to be learned concerning the free changes leading to vascular re-arrangement, such as the role played by endothelium and cytokines, in intestinal schistosomiasis. However repair seems to be processed within a few months in the murine model, being completed by the vascularization of the involuting granulomas and the straightening-out of the submucosal veins. REFERENCES Andrade ZA, Brito PA 1981. Evolution of schistosoreal hepatic vascular lesions after specific chemotherapy. Am J Trop Med Hyg 30: 1223-1227. Andrade ZA, Silveira GM 1990. Esquistossomose Intestinal. Gastroenterologia Etwloscopia Digestiva 9: 158-185. Andrade ZA, Andrade SG, Susin M 1974. Pathological changes due to massive schistosomal infection in nmn. Revlnst Med trop S Paulo 16: 171-177. Cheever AW 1969. Quantitative comparison of the intensity of Schistosoma mansoni infections in man and in experimental animals. Trans R Soc Trop Med Hyg 63: 781-795. Cheever AW 1970. Relative resistance of the eggs of human schistosomes to digestion in potassium hydroxide. Bull WHO 43: 601-603. Cheever AW & Andrade ZA 1967. Pathological lesions associated with Schistosoma mansoni infection in man. Trans R Soc Trop Med Hyg 61: 626-639. Kloetzel K 1971. Tissue reaction to Schistosoma mansoni ova. III - Micro-circulation in the intestine of infected mice, as studied by injection of contrast material. Rev Inst Med trop Sao Paulo 13: 51-56. Nagy BA, File SK, Smith JH 1981. Changes in the enteric vasculature of mice infected with Schistosoma mansoni. Am J Trop Med Hyg 30: 999-1009.
Santos RO, Baeoosa JR APt, Andrade ZA 1992. Dynamics of fibrosis production and resorption in intestinal schistosomiasis of mice. Mem Inst Oswaldo Cruz 87:25-31. Weinstock JV, Boros DL 1981. Heterogeueity of the granulomatous response in the liver, colon, ileum, and ileal Peyer's patches to schistosome eggs in murine schistosomiasis mansoni. J Immunol 127: 1906-1909. Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95078a.gif] [oc95078b.gif] [oc95078c.gif]Photo images[oc95078b.jpg] [oc95078d.jpg] [oc95078a.jpg] [oc95078c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}