 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Interaction between Poultry and Triatoma infestans Klug, 1834 (Hemiptera: Reduviidae) in an Experimental Model Nicolas J Schweigmann, Silvia Pietrokovsky, Osvaldo Conti, Victoria Bottazzi, Delmi Canale*, Cristina Wisnivesky-Colli Unidad Ecologia de Resetvorios y Vectores de Parasitos, Depaflamento de Ciencias Biol6gicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 1428, Buenos Aires, Argentina *Servicio Nacional de Chagas, Cdrdoba, Argentina
Code Number: OC95084
Size of Files:
Text: 14K
Graphics: Line Drawings (gif) 10K
This paper compares the predation pressure that ducks
and chickens exert on triatomines. For the tests, these birds
were placed in individual boxes together with a known number
of Triatoma infestaris and left to interact from 6 p.m
till the next morning, involving a long lasting period of
complete darkness limited by two short-term periods of
semi-darkness. There was a shelter which could prevent the
bugs from being predated The number of live and dead
triatomines was recorded considering missing bugs as predated
by the birds. Ducks exhibited a greater predatory activity
than chickens, that could be due to a long term active period
at night while chickens sleep motionless from sunset to dawn.
Surviving triatomines that had fed on chickens outnumbered
those fed on ducks suggesting that these were less accessible
to the triatomine biting.If ducks are better than chickens to detect and eat bugs and to interfere with their feeding in the field, an increase in duck number might help to diminish triatomine density. Further research is needed to determine the feasibility of application of these experimental results. Key words: Triatoma infestans - ducks - poultry - Chagas disease control - Argentina In the rural areas of Santiago del Estero, Argentira, where Chagas disease is endemic, chickens (Gallus gallus) and ducks (Anas sp.) wander around the peridomiciles pecking at grains and insects.
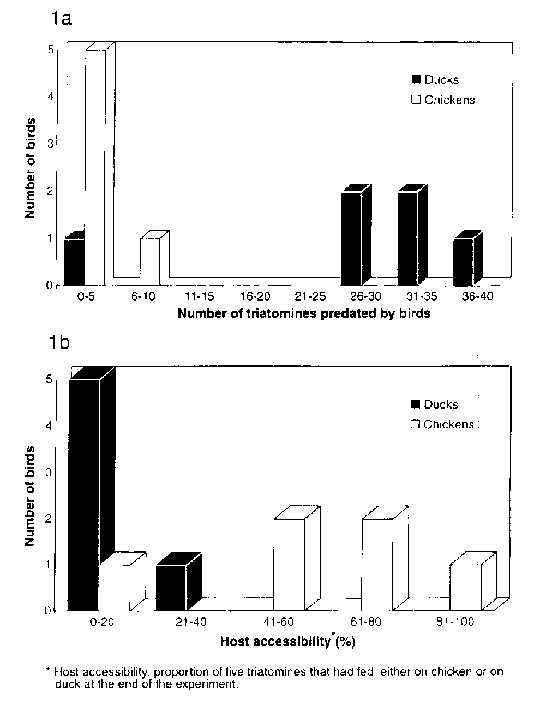
Schofield (1985) suggested that chickens would exert a considerable predation pressure on the vector insect, Triatoma infestans Klug, 1834. However, this predation impact could be decreased by the fact that - unlike triatomines chickens are diurnal animals (Rossell 1976). Moreover, chickens help keeping stable colonies of T. infestans since they are usually selected as a blood source by these triatomines (WisniveskyColli 1987). Besides, Gajate et al. (1994) demonstrated a close association between chickens and the presence of T. infestans in peridomestic ecotopes. On the other hand, ducks are less abundant than chickens because they are seldom reared in the area, and up to now their relationship with triatomines has never been studied. After several years of field work we observed a different behaviour in chickens and ducks: while chickens sleep motionless from sunset to dawn, ducks alternate motionless and active periods. This led us to wonder whether ducks would be more efficient predators of triatomines than chickens and as good hosts as them. In this paper we analyse the bird-triatomine interaction under experimental conditions to compare the eventual negative effect these birds would cause in the abundance of T. infestans populations. MATERIALS AND METHODS The experiment was-carried out in a field laboratory in the village of Amama, Departamento Moreno, Provincia de Santiago del Estero, Argentina, in March, 1991. Panama had been sprayed five years before, thus precluding any residual effect of the insecticide. The experimental device consisted of a hardboard box (50 x 50 x 45 cm) - where the birds could move freely - with a transparent sliding lid at the top. There was a removable wooden panel (40 x 14 cm) on one of the walls, which acted as a shelter for triatomines; it was placed 5 cm above the floor and 0.5 cm from the wall, leaving its lateral and lower edges open. A window (10 x 10 cm) covered with a fine wire net provided ventilation and a thin layer of sand Was spread on the floor to absorb humidity from the excreta. The third and fifth instar nymphs of T. infestans used in the trials were provided by the insectary of the Servicio Nacional de Chagas, while the adult bugs were supplied by local inhabitants. All triatomines had been starved for 25 days. Chickens and ducks were borrowed locally since we wanted birds accostomed to peck insects in the field. Fowl was not under veterinary treatment of any kind. Both birds and insects were used only once. One chicken and one duck were tested simultaneonsly in separate boxes (two cases per night) and they were left in a room where light could get in through windows. A total of eight trials including 280 insects were perfomaed for both chickens and ducks: six of them involved 40 nymphs each, and the other two involved 20 adults of T. infestans each. The experiment proceeded as follows: bugs were introduced in the box at 6 p.m. - one hour before the bird - and they were left until the next morning. Approximately at 7 a.m., the birds were removed from the cages and examined carefully as well as the boxes. Therefore, the experiment was carried out during a long lasting period of complete darkness limited by two short-term periods of semi-darkness. Live and dead triatomines were reckoned up considering missing bugs as predated by the bird. We used Montenegro's (1983) and Rossell Reyes' (1984) methods - for adults and nymphs respectively - to determine if the bug had fed or not on the bird. Any imago showing blood in the promesenteron and nymph with a slightly convex abdomen were considered as fed. Host accessibility was defined as the bird acceptance to the triatomine biting and it was estimated as the proportion of live triatomines that had fed either on chickens or on ducks at the end of the experiment. The x^2 test for proportions was used to compare the predatory activity, and host accessibility was analysed by the Wilcoxon test (Sokal & Rohlf 1969). RESULTS Predation - Dead nymphs were found in four trials with chickens and in three with ducks, and in most cases the number of dead insects ranged from 1-3. In one trial, though the chicken did not eat any nymph, 28 dead bugs had been trampled on. Considering the six trials involving nymphs (Fig. la), ducks predated between 26-37 nymphs each (>65% of the initial number) in five cases, and one duck ate only one insect. Conversely, three chickens did not eat nymphs and the remainder predated 2-8 (<20%). A similar result was observed in the trials where 20 adult bugs were employed: ducks ate almost all the insects (17-19/20) while the chicken just ate 2-5/20 bugs (not shown in figures). TABLE Global comparison of host accessibility of chickens and ducks
-------------------------------------------------------------
Host accessibility a %
Chicken Ducks
-------------------------------------------------------------
Triatoma infestans
Nymphs b 119/192 18/71
62% 25%
Adults 24/33 1/4
73% 25%
-------------------------------------------------------------
a: host accessibility = proportion of live triatomines that
had fed either on chicken or on duck at the end of the
experimentb: p < 0.001
Host accessibility - Fig. 1b shows the rate of live nymphs fed either on chickens or on ducks. It can be observed that chickens allowed a higher number of nymphs to feed on them. Table shows that overall host accessibility of ducks (25%) for nymphs was significantly lower than that found in chickens (62%) (p<0.001). A similar trend can be seen when adults bugs are considered. DISCUSSION In the dwellings of endemic areas, bugs come out from their hiding places at night to bite sleeping humans and domestic animals. This behaviour that had already been described by Carlos Chagas in 1909, was experimentally confirmed by Lazzari (1992) who demonstrated the existence of two peaks of locomotor activity in T. infestans: one at sunset related to food search and another at dawn when insects are looking for shelter. Schofield (1985) suggested that bugs would feed at night to avoid predation by hosts of diurnal habits such as chickens. Chickens do not show spontaneous movements during the deep sleeping period (Ookawa Gotoh 1965). On the contrary the noctumal activity of some anatids could be demonstrated due to the recent development of telemetry. This method showed that, for instance, the blue duck (Hymenolaimus malacorhynchos) was active not only at daytime, but at sunset, at midnight and before dawn too (Douglas & Pickard 1992). The high predation performed by ducks in our experiments could be explained by their long term nocturnal period of activity; the pecking heard in the boxes containing ducks during total darkness would denote this fact. Jiron and Zeledon (1982) pointed out the importance of host response to bug feeding. These authors studied the feeding performances of nymphs of T. infestans and other triatomines in an experimental device using four hosts: dog, opossum, hen and toad, during 2 hr at midday and 14 hr at night. They found that a significantly higher proportion of triatomines had fed on those hosts that were resting (dog and hen during the night, opossum in daytime). Therefore, host activity could be related to a higher sensibility to triatomine biting. Considering host accessibility as an indirect indicator of host sensibility, we experimentally found that ducks would be more irritable than chickens. Summing up, the combination of these two factors, an outstanding predation aptitude and a low tolerance to triatomine biting characterize ducks as a non-profitable blood source for triatomines. In the field, ducks probably are more skillful than chickens to detect and eat unsheltered bugs as well as to interfere with the fullfilness of the blood meal of the triatomines they attract. Perhaps an increase in the number of ducks might diminish the density of intradomicile and peridomestic triatomines, particularly after spraying the dwellings with insecticides. Further research is needed to determine whether ducks should be considered as an important tool in the biological control of vectors in the field. ACKNOWLEDGEMENTS To the Citatti family for providing local facilities for our field laboratory, to the Moyanos for hostage and for lending us the birds and to Bea Ferrer for her kind assistance with the English version of the manuscript. REFERENCES: Chagas C 1909. Nova tripanozomiaze humana. Estndos sobre a moffologia e ciclo evolutivo do Schizowypanum cruzi n.gen., n. sp., ajente etiologico de nova morbida do homen. Mem Inst Oswaldo Cruz 1: 159-218. Douglas ME, Ross Pickard C 1992. Telemetry of body tilt for automatic data logging of blue duck diel hehaviour, p. 599-611. In IM Priede, SM Swift (eds). Wildlife telemetry. Remote monitoring tracking of animals. Elis Hotwood. Chichester, England. Gajate P, Bottazzi V, Pietrokovsky S, WisniveskyCoili, C 1994. Relaqao entre O Triatoma infestans, as aves domesticas e os homeus, num povoado de Santiago del Estero, Argentina. Cad Saade Publ 10: 53-57. Jiron LF, Zeledon R 1982. Preferencias alimentarias de tres especies de Triatominae (Hetniptera: Reduviidae) en condiciones experimentales. Rev Biol Trop 30: 151-159. Lazzari CR 1992. Circadian organization of locomotion activity in the haematophagous bug T. infestans. J Insect Physiol 38: 895-903. Montenegro SC 1983. Detetminacion de reservas alimentarias en T. infestans, Klug. 1834 (Hemiptera: Reduviidae) en base a caracteres extemos. Adultos. Physis (secckin C) 4: 159-167. Ookawa T, Gotoh J 1965. Electroencephalogram of the chicken recorded from the skull under various conditious. J Comp Neur 124: 1-14. Rossell OJ 1976. Evaluacion de la transmisidn de la enfermedad de Chagas en dos caserios del Estado Guarico, Venezuela, sometidos a rociamientos. Trabajo especial pard optar a la categoria de profesor Asistente, Facultad de Ciencias, Universidad de los Andes, Merida, Venezuela, 76 pp. Rossell Reyes O 1984. A comparison of the feeding perromances of Rhodnius prolixos Stal and Rhodnius robusstus Larroasse (Hemiptera: Triatominae). Doctoral Thesis, Univevsity of London, 339 pp. Schofield CJ 1985. Population dynamics and control of Triatoma infestans. Ann Soc Beige Med Trop 65 (Suppl. 1): 149-164. Sokal RR, Rohlf FJ 1969. Biometry. WH Freeman, San Francisco. U.S.A. xxi+ 776 pp. Wisnivesky-Colli C 1987. Feeding patterns of Triatominae in relation to transmission of American Tpypanosomiasis, p. 99-117. In A Stoka, R Brenner (eds). Chagas'disease Vectors, Vol.l, CRC Press Inc, USA. Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc95084a.gif] |
| |||||||||

{kind=link}