 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ultrastructure of the Male Terminalia of Cochliomyia hominivorax and C. macellaria Antonio Cesar Rios Leite Departamento de Parasitologia, Instituto de CiEncias Biologicas da Universidade Federal de Minas Gerais, Caixa Postal 486, 31270-901 Belo Horizonte, MG, Brasil
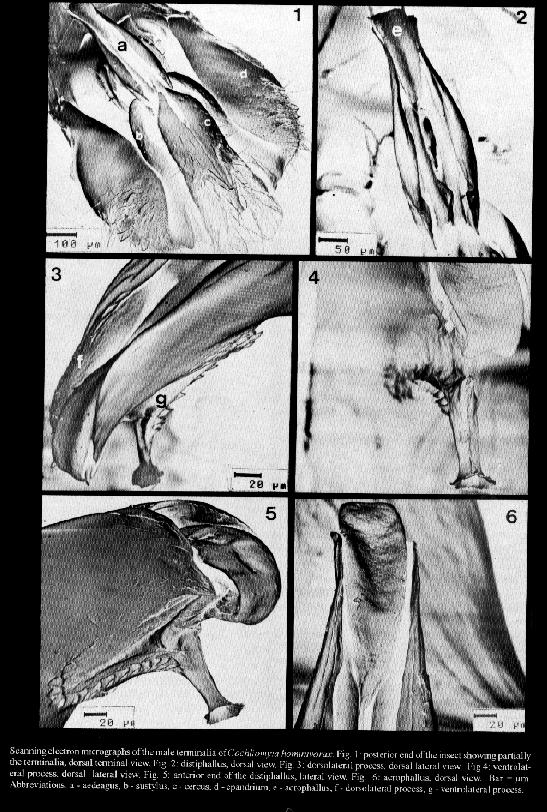
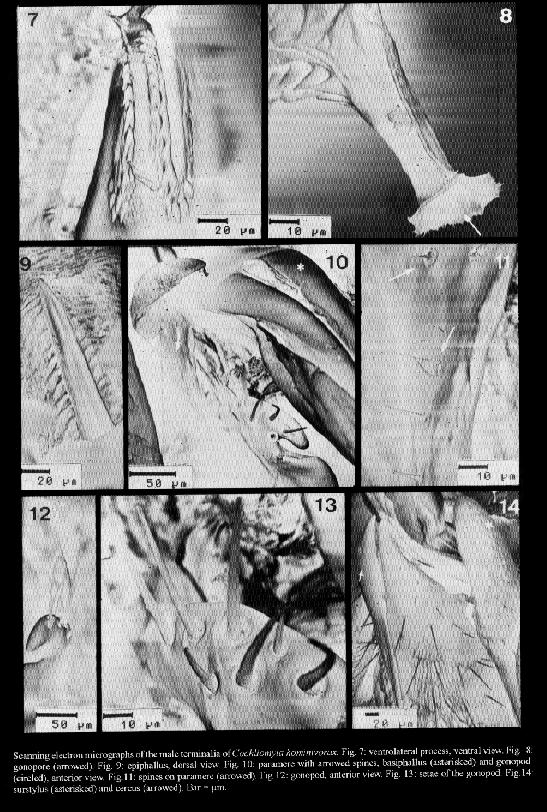
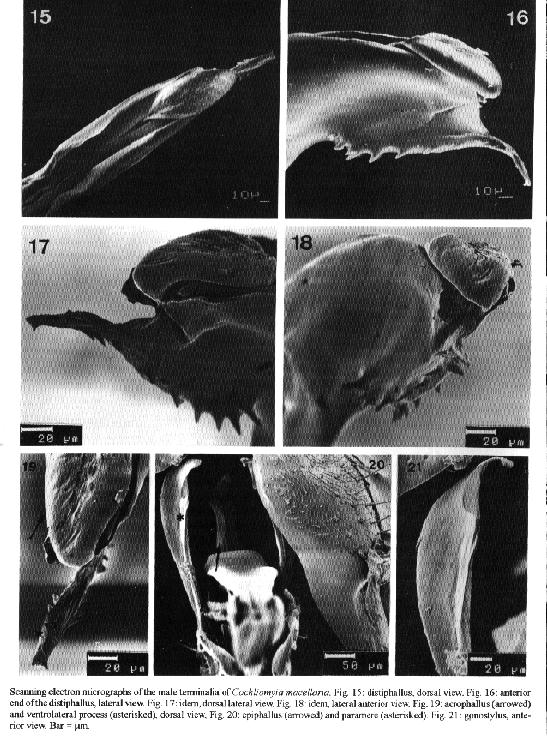
Cochliomyia hominivorax and C. macellaria are species that cause primary and secondary myiasis, respectively, of medical and veterinary importance in the New World. The male terminalia of both insects show morphological differences that are now described from scanning electron microscope (SEM) observations. Key words: Diptera - Calliphoridae - Cochliomyia hominivorax - Cochliomyia macellaria - terminalia - ultrastructure The screwworms, Cochliomyia hominivorax (Coquerel) and C. macellaria (Fabricius), are indigenous flies of the New World (James 1970). C. hominivorax was introduced into the Old World, in Libya (Gabaj et al. 1989) but has recently been eradicated (Lindquist et al. 1992). C. hominivorax is responsible for primary cutaneous myiasis in mammals, including man, causing the severe economic loss in livestock in the Neotropical Region. C. macellaria causes secondary myiasis in the same hosts (Snow et al. 1981, Miller 1982, Baumgartner & Greenberg 1983, Guimaraes et al. 1983). The male terminalia of both species of screwworm flies was described under light microscope (LM) ( Cushing & Patton 1933, Laake et al. 1936, Mazza & Jorg 1939, James 1947, Hall 1948, Guimaraes et al. 1983). In C. hominivorax, different strains show small morphological distinctions in the male genitalia (Baumgartner & Greenberg 1983). Using SEM, physical changes caused by mating have been observed in the male genitalia of C. hominivorax (Gagne & Peterson 1982). Ultrastructural descriptions of the aedeagal apodeme of C. hominivorax has been used to determine the fly's age (Ellison & Hampton 1982). Richardson et al. (1982a) reported genetic diversity in populations of C. homonivorax by means of caryotypic studies, which were associated with ultrastructural differences in the male genitalia. Continuing the studies of Leite and Guevara (1993), the male terminalia of C. hominivorax and C. macellaria are described herein based on tridimention observations. MATERIALS AND METHODS The third instar larvae of C. hominivorax and C. macellaria were collected from the field, in Belo Horizonte, Minas Gerais, Brazil (19 55' S, 43 56' W, alt. 852 m) and adult specimens were reared in the laboratory (Guevara 1986). The terminalia dissected from virgin males of the flies were processed for SEM, based on the method described by Leite and Williams (1988). The specimens were dried at room temperature, submitted to critical point drying, spattered with gold, and studied by scanning electron microscopy. The nomenclature of the male terminalia is that of McAlpine (l981). RESULTS Cochliomyia hominivorax The elements of the male terminalia are partially illustrated in Fig. 1. The aedeagus is arched and formed by a solid basiphallus and a ornamented distiphallus. The latter bears internally two forcep-like arms from the basiphallus (Fig. 2), and, laterally, the distiphallus has a rigid plate, the dorsolateral process, wrinkled internally and recurved inward at the dorso-mediate region (Fig. 3). The acrophallus has a tongue-like membranous structure (Figs 3-6), which enters the dorsal anterior union of the distiphallus arms and is situated between the two lateral plates. At the apex, the acrophallus is recurved ventrally and passes between two pointed spine-like processes. The ventrolateral process (Figs 4, 5, 7) has several strong thorn-like spines, and a free anterior end with an opening, the terminal gonopore (Fig. 8). The epiphallus is slender and distally pointed (Fig. 9). The paramere (Fig. 10) bears a strong gonocoxite, with long and pointed spines and a twisted gonostylus with short small spines (Fig. 11). The plate-like gonopod bears 6-10 short and long setae and a subapical groove (Figs 12-13). The robust surstylus (Fig. 14) is spatulate and a slightly recurved distally. The broad short cerci (Fig. 14) are united at the base and separated distally by two small apically acuminate arms. Scanning electron micrographs of the male terminalia of Cochliomyia hominivorax. Cochliomyia macellaria The aedeagus bears a strong basiphallus and an ornamented distiphallus (Fig. 15) formed by two forcep-like arms from the basiphallus. The dorsolateral process of the distiphallus is arched dorsally and fused anteriorly. The acrophallus has a membrane-like structure, beret-shaped (Figs 16 -19), situated dorso-terminally on the lateral plates: its anterior end is free and its posterior end penetrates the distiphallus. The ventrolateral process (Figs 16-19) has a few strong spines, and a long terminal projection. The terminal gonopore, at the apex of the terminal projection, could not be examined because it was not open. The epiphallus (Fig. 20) is broad and apically slender. The paramere (Fig. 20) shows a stout gonocoxite with large setae and several spines (Fig. 23), and a gonostylus with spines and a latero-apical fold (Figs 21-22). The gonopod has four setae and a asymmetric groove (Fig. 24). The surstylus is robust and slightly twisted at the anterior end (Fig. 25). The long cerci, united at the base, are separated distally by two long arms (Figs 25-26). DISCUSSION The taxonomic distinctions between adults of C. hominivorax and C. macellaria are mainly based on descriptions of the male and female terminalia (Cushing & Patton 1933). By LM, the morphological differences between the two species of screwworms can be particularly identified: in C. hominivorax, the aedeagus supports an acrophallus with a ventrolateral process showing a short projection, the gonopod bears many setae and the cerci show short arms whereas C. macellaria has a ventrolateral process with a long terminal projection, the gonopod with a small number of the setae and the cercus with long arms (Cushing & Patton 1933, Mazza & Jorg 1939, James 1947, Guimaraes et al. 1983, Shewell 1987). In C. hominivorax, from different strains, small variations in the male genital morphology were found, under LM, in the epiphallus angle, in the forms of the distal end of the aedeagus, paramere and apodeme (Braumgartner & Greenberg 1983). Indeed, the forms and lengths of the apodemes and distal end of the aedeagus show distinctions, but the slight differences admited and drawn by the same authors, for the other genital elements, are very difficult to define at optical level. Based on SEM, Richardson et al. (1982a) supplemented their studies concerning nine genetic types of C. hominivorax populations, and concluded that four types had structural differences in the male genitalia. The four types have characters that are not seen in C. hominivorax from Belo Horizonte: type 009 (factory-reared strain, from Arizona and New Mexico) shows more short setae on the gonopod, and the paramere with a fold and a pointed apex. Such folds, although in different positions, and a pointed apex were also observed in the other reared flies: type A (from New Mexico, Arizona and California), type B ( from New Mexico and California) and type F (from New Mexico). The ultrastructural diferences in the epiphallus, gonopore and paramere of C. hominivorax caused by attempts to mate or after mating, have been reported (Gagne & Peterson 1982). The genitalia of the male from Florida that had mated is similar to that seen in the present work because it has seven setae on the gonopod and a fold on the paramere. This last characteristic was also observed in mated specimens from Mexico. The position of the epiphallus angle represents a caracter of distinction between C. hominivorax strains (Baumgartner & Greenberg 1983), signifies changes in the mated or unmated flies (Gagne & Peterson 1982), and may be a consequence of injury caused during handling or preparation of the terminalia. The results published by Gagne and Peterson (1982) were considered ambiguous by Richardson et al. (1982b). The latter compared the genitalia type F, with 1-day-old (corresponding unmated fly) and that of genitalia type B, with 8-day-old (from mated fly), and these were similar to that of the mated male and unmated male, respectively, as described by Gagne and Peterson (1982). The thorn-like spines on the ventrolateral process described herein do not differ from those figured by Gagne and Peterson (1982). The short spines on the paramere are perhaps visible in figure B, published by Richardson et al. (1982b). Although the differences between the terminalia of the C. hominivorax and C. macellaria are easily identifed under LM, such differences added to the others described in this work, based on tridimensional study, represent a more realistic and detailed description of these two important Calliphoridae, and may contribute towards an improvement in morphological knowledge of screwworms. REFERENCES Baumgartner DL, Greenberg B 1983. The primary screwworm fly, Cochliomyia hominivorax (Coquerel) (Diptera, Calliphoridae), in Peru. Rev Bras Biol 43: 215-221. Cushing EC, Patton WS 1933. Studies on the higher Diptera of medical and veterinary importance. Cochliomyia americana sp. nov., the screw-worm fly of the New World. Ann Trop Med Parasitol 27: 539-551. Ellison JR, Hampton EN 1982. Age determination using the apodeme structure in the adult screwworm flies (Cochliomyia hominivorax). J Insect Physiol 28: 731-736. Gabaj MM, Wyatt NP, Pont AC, Beesley WN, Awan MAQ, Gusbi AM, Benhaj KM 1989. The screwworm fly in Libya: a threat to the livestock industry of the Old World. Vet Rec 125: 347-349. Gagne RJ, Peterson RD 1982. Physical changes in the genitalia of males of the screwworm, Cochliomyia hominivorax (Diptera: Calliphoridae), caused by mating. Ann Entomol Soc Am 75: 574-578. Guevara JDE 1986. Biologia de duas amostras brasileiras de Cochliomyia hominivorax (Diptera: Calliphoridae) submetidas ou nao a irradiacao gama. MSc. Thesis. Universidade Federal de Minas Gerais, Belo Horizonte, 49 pp. Guimaraes JH, Papavero N, Prado AP 1983. As miiases na regiao neotropical (identificacao, biologia, bibliografia); familia Calliphoridae (gEnero Cochliomyia). Rev Bras Zool 1: 270-335. Hall DG 1948. The blow flies of North America, Vol. 4. Thomas Say Foundation. Entomological Society of America, College Park, Maryland, 477 pp. James MT 1947. The flies that cause myiasis in man. US Dept Agric Misc Publ 631, Washington, 175 pp. James MT 1970. Family Calliphoridae. A catalogue of the Diptera of the Americas South of the United States. Museu de Zoologia da Universidade de Sao Paulo, Sao Paulo, 102.1-102.28 pp. Laake EW, Cushing EC, Parish HE 1936. Biology of the primary screw worm fly, Cochliomyia hominivorax, and a comparasion of its stages with those of C. macellaria. US Dept Agric Tech Bull 500, Washington, 24 pp. Leite ACR, Williams P 1988. Redescricao de adultos de Metacuterebra apicalis (Diptera: Cuterebridae). Mem Inst Oswaldo Cruz 83: 493-507. Leite ACR, Guevara JDE 1993. Scanning electron microscopy of the larval instars of Cochliomyia hominivorax. Med Vet Entomol 7: 263-270. Lindquist DA, Abusowa M, Hall MJR 1992. The New World screwworm fly in Libya: a review of its introduction and eradication. Med Vet Entomol 6: 2-8. Mazza S, Jorg ME 1939. Cochliomyia hominivorax (Coq.) americana. C. y P., estudio de sus larvas y consideraciones sobre miasis. Investigaciones sobre dipteros argentinos. I. Miasis. p. 3-46. Mision de Estudios de Patologia Regional Argentina, Pub 41, Buenos Aires. McAlpine JF 1981. Morphology and terminology-adults, Mon 27, p. 9-64. In JF McAlpine et al. Manual of Nearctic Diptera. Research Branch, Agriculture Canada, Ottawa. Miller R 1982. Screwworm, ser c, v 1, p. 263-267. In H Steele, C.R.C Handbook. Series in Zoonoses. Parasitic Zoonose. C.R.C. Press, Boca Raton. Richardson RH, Ellison JR, Averhoff WW 1982a. Autocidal control of screwworms in North America. Science 215: 361-370. Richardson RH, Ellison JR, Averhoff WW 1982b. Mating in screwworm populations? Science 218: 1142-1145. Shewell GE 1987. Calliphoridae, Mon 28, p. 1133-1145. In JF McAlpine et al., Manual of Nearctic Diptera Research Branch, Agriculture Canada, Ottawa. Snow JW, Siebenaler AJ, Newell FG 1981. Annotated bibliography of the screwworm, Cochliomyia hominivorax (Coquerel). US Dept Agic Sci Adm, Agric Rev Man, South Ser 14, New Orleans, 32 pp.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95093c.gif] [oc95093a.gif] [oc95093b.gif]Photo images[oc95093b.jpg] [oc95093c.jpg] [oc95093a.jpg]Line drawing images[oc95093d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}