 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Hemagglutinating and Fusogenic Activities of Newcastle Disease Virus: Studies on Receptor Binding Specificity and pH-induced Conformational Changes ESS Couceiro/+, |NSS Couceiro*, MC Cabral Departamento de Virologia, Instituto de Microbiologia, CCS-UFRJ Cidade Universitaria, 21941-590 Rio de Janeiro, RJ, Brasil
Code Number: OC95102
Size of Files:
Text: 25K
Graphics: Line Drawings (gif) 20K
Photos (jpg) 31K / Halftones (gif) 33K
Vaccinal and wild strains of Newcastle Disease virus (NDV) were analyzed for cell receptor binding and fusogenic biological properties associated with their HN (hemagglutimn-neuraminidase) and F (fusion protein) surface structures respectively. The evaluation of the biological activities of HN and F was carried out respectively by deterntination of hemagglutinating titers and hemolysis percentages, using erythrocytes from various animal origins at different pH values. Significant differences in hemag- glutination titersfor some strains of NDV were detected, when interacting with goose, sheep, guineapig and human "O" group erythrocytes at neutral pH. Diversity of hemolysis percentages was observed between different NDV strains at acid pH. These analysis were developed to evaluate particular aspects of the actual influence of the receptor specificity and pH on the receptor binding and fusogenic processes of Newcastle Disease viruses. Key words: Newcastle Disease virus - hemagglutinin - neuraminidase - fusion protein - variants Newcastle Disease viruses (NDV) are RNA viruses that present glycoproteins inserted in theft envelope structure, playing important roles on different steps of the infectious process. These vimses exhibit HN (hemagglutinin-neuraminidase) and F (fusion protein) glycoproteic spikes as their surface structures which are responsible for cell receptor binding (HN), cleavage of cell receptors (HN) and fusion (F) processes, dimng the virus replication cycle (Marsh & Helenius 1989, Marsh & Pelchen Matthews 1993, Vainiopaa & Hyypia 1994). HN structures of NDV which exhibit receptorbinding activity on gangliosides during the viruscell adsorption process, have been studied by hemagglutination test. In analysis nslrqg influenzavirus as model, the amount and quality of sialic acids receptor residues which vary among erthrocytes from different animal origins, show influence on the viruscell receptor bincling process, where the crucial conformational changes of HA structures are pH-dependent CMarkwell et al. 1986, Katz & Webster 1988, Phyalla et al. 1988, Marsh & Helenins 1989, Marsh & pelchen-Matthews 19 9 3 ). This work was supported by FINEP, FAPERJ, CNPq and CAPES +Corresponding author. Present address: Biomanguinhos-Funda~ao Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil Received 22 September 1994 Accepted 10 February1995 F glycoproteins, which are responsible for the fusion process, exhibit biological activities after cleavage to F1 and F2 enzymatic activity (eg. trypsin). In this process, pH-independent fusion occurs after insertion of the hydrophobic moiety of F1 into the target membrane, which is in vitro revealed by hemolysis reaction, when erythrocytes are used as cellular model (Marsh & Helemus 1989, Marsh & Pelchen-Matthews 1993, Stegmann 1994). Analysis on the influence of cell receptors and pH on the adsorption and hemolysis processes have been developed, using non purified or non standardized preparations of viruses from different families (Clark & Nagler 1943, Winslow et al. 1943, Muller & Stanley 1944, Chu 1948, Clavell & Bratt 1972). The aim of this study was to analyze different purified and standardized NDV strains for particular aspects of their hemagglutinating and hemolytic activities, using different parameters. The pH was analyzed as interfering factor on hemagglntination and fusion processes.
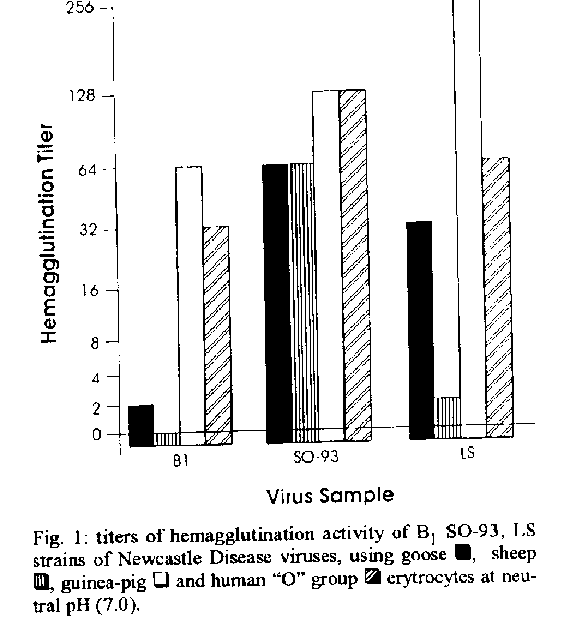
MATERIALS AND METHODS Viruses - Lentogenic (B1 and La Sota) and velogeinc (S0-93)* virus samples were inoculated into the allantoic cavity of 10 day-old chicken embryonated eggs and incubated for 48 hr at 37 C. *Wild sample isolated in Brazil which was provided by Dr Ary Monteiro de Souza, Dept de Virologia, Empresa Brasileira de Agropecuafia, RJ. The allantoic fluids were harvested and clarified by centrifugation at 3.500 x g for 30 min at 4 C. The virus samples were 50 x concentrated (rediluted in Phosphate Buffer Solution-PBS pH 7.0 after ultracentrifugation at 80.000 x g for 60 min), purified in 40% potassium tartrate 5% glycerol continous density gradient by ultracentrifugation at 100.000 x g for 120 min. The virus bands were collected, diluted (1:10) in PBS pH 7.0 with 2mM EDTA and centrifuged at 4 C again for 60 min at 80.000 x g. The pellets were reddiluted using PBS- EDTA and stored at -20 C until utilization (Couceiro etal. 1994). Evaluation of the protein content of the virus samples was processed by the Bradford method (Bradford 1976) for the obtainment of standardized concentrations of virus proteins to develop hemagglutination and hemolysis assays. Erythrocytes - Human "O" group (Rh+), guinea-pig, sheep and goose erythrocyles were used. Eyrthrocytes were collected in Alsever solution and stored at 4 C (Pinto et al. 1994). After washing for three times, they were adjusted at 10% concentration (v/v) in saline (0.15 M NaCl). Afterwards those 10% erythrocyte suspensions were diluted in adequate buffer for each test (Pinto et al. 1994). Hemagglutination test - Virus samples were diluted serially in equal volumes (25 ul) for saline (0.15 M NaCI) and to each dilution of virus was added 25 ul of 1% mammalian or 0.5% goose erythrocyte suspension at pH values ranging from 5.0 to 8.6. Mammalian and goose erythrocyte snspensions at pHvalues ranging from 5.0 to 8.6 were prepared in Michaelis barbital buffer (Pinto et al. 1994). Each assay was developed in triplicates for twice, the geometrical average of the ti- ters representing its final titer. The titer of each assay was reported as the reciprocal of the highest virus dilution responsible for complete hemagglutination at 4 C (Pinto et al. 1994). Hemolysis test (Lenard & Miller 1981) Hemolysis activity of NDV samples was determined in a total volume of 1.5 ml in capped tubes, containing 0.1 ml of human "O" group (Rh+) erythrocyle suspensions (10%), 0.1 ml of each virus sample at two standardized protein concentrations (1 and 5 ug/ml) and 1.3 ml of Sorensen's phosphate buffer (Pinto et al. 1994) at different pH values. The capped tubes were incubated for 60 min in cold bath (0-4 C), which was followed by incubation for 60 min at 37 C with frequent mixing. After incubation, the tubes were centrifuged for 10 min at 600 x g and the amount of released hemoglobin (Hb) in the supernatant was estimated by absorbance (Ab) measurement, using a Perkin Elmer spectrophotometer at 590 nm. Each assay was developed in triplicates for twice, considering the arithmetical average of the data (Ab) as the final result of each type of experiment. The hemolysis level of each assay (Pinto et al. 1994) was determined by comparison of the measured Ab with that of 1.0 ml of erythrocyte suspension in 1.4 ml of buffer solution (spontaneous hemolysis) and that of 1.0 ml of erythrocyte suspension in 1.4 ml of deionized water (total hemolysis). Evaluation of different NDV samples for he- magglutinating activity at neutral pH, using erythrocytesfrom different animal origins - The virus samples were diluted in PBS with posterior addition of erythrocyte (mammalian or goose) suspensions at neutral pH (7.0). The technical procedures for hemagglutination were those described above. Evaluation of NDV samples for hemolysis activity at acid pH -The NDV samples were diluted to 1 ug/ml protein concentration in PBS and they were analyzed by hemolysis test, which was developed such as previously described, with utili- zation of erythrocytes in Sorensen's phosphate buffer (pH 5.0). Erythrocytes in deionized water (total hemolysis) and erythrocytes at the different pH values (spontaneous hemolysis) were used as controls. Evaluation of B1 strain of NDV for hemolysis activity after variation of pH (5.0 - 6.0) - The sample (5 ug/ml protein concentration) showing the lowest hemolysis percentage at pH 5.0 was exposed at pH variation, using 1N NaOH, as indi- cated below: pH variation: 5.0 - 6.0 pH 5.0 - pH 6.0 (for 5 min) NaOH Evaluation of the hemagglutinating activity at large pH range, using that B1 strain of NDV that exhibits the lowest hemagglutinating titers at neutral pH - The sample was diluted in PBS with posterior addition oferythrocyte (sheep or goose) snspensions at pH values ranging from 5.0 to 8.6, which were prepared in Michaeli's bafoital buffer (Pinto et al. 1994). The technical procedures for hemagglutination were those described above. Analysis of that B1 strain of NDV analized by hemagglutination and hemolysis experiments, using electron microscopy - NDV virus particles of that purified B1 strain used in experiments were observed by electron microscopy, showing the influence of neutral pH, concentration and purification processes on the original virus structures. A negative combined (4 % phosphotungstic acid and uranyl acetate for one min each one) staining technique was developed as described by Fonseca et al. (1984) using a Phillips 301 transmission electron microscope. RESULTS AND DISCUSSION NDV preparations were analyzed for their he- magglutinating and hemolytic activities, using cells from different origins as targets, while ambient pH was used as interfering factor on these processes. A variation in the hemagglutinating activity of NDV (B1, La Sota and S0-93 strains) at neutral pH could be observed by the utilization of goose, sheep, guinea-pig and human "O" group erythrocytes (Fig. 1), that might be suggestive of the existence of different subpopulations or variants of virus particles in the same virus preparation. Variants with higher affinity for determined cell surface structures, which could be selected during the virus propagation process. Influenza viruses show a known standard behavior due to the importance of the receptor specificity, during the hemagglutinin-cell receptor interaction process. In parallel experiment, A/PR/8/ 34, sample of influenza viruses exhibited a very significative variation of hemagglutinating titers (from 256 to 4096) due to the presence of specific cell receptors on membranes of avian and mammalian erythrocytes (ESS Couceiro, JNSS Couceiro & MC Cabral. unpublished results). The propagation of that sample in embryonated chicken eggs selected a virus preparation with higher affinity to sialic acid alpha-2,3Gal-containing receptors (Rogers & D'Souza 1989) which are found in higher percentage on avian (goose) cells (hemagglntinating titer equal to 4096). Lower hemagglutinating titers were observed to mammalian cells as sheep (512), guinea-pig (1024) and human
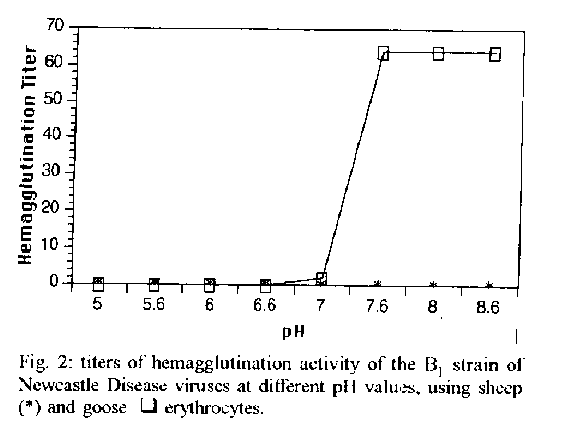
However the same level of variation was not observed when paramyxoviruses were utilized as model. The variation was not significant for the SO-93 sample of NDV (titers from 64 to 128) as can be observed in Fig. 1, when erythrocytes from different animal origins were used. The results present the preparations of Bland La Sota lentogenic samples of NDV as the most heterogeneous mixtures of paramyxovirus variants (hemagglutinating titers varying from <2 to 256), also exhibiting higher hemagglutinating titers than that SO93 velogenic sample. The cell receptors for pammyxoviruses (NDV) are expressed by different complex gangliosidic structures as GDIa, GTIb and GQIb (Markwell et al. 1986, Haywood 1994), which could be more plentiful on cell surfaces of erythrocytes originated from specific animal origins. The results from hemagglutination assays obtained with sheep, goose, guinea-pig and human "O" group erythrocytes showed the occurrence of variability in the amount and specificity of erythrocyles receptors for paramyxovirus, as described by Martoneetal. (1973). Thus, it can be considered that the majority of the NDV samples presented virus particles that were able to recog- nize more efficiently those cellular gangliosidic receptors exhibited on guinea-pig and human "O" group erythrocytes better than on those from other origins. 60% homology level at amino acid level between different influenza virus strains for receptor binding HN structures could also explained these differences (Vainionpaa & Hyypia 1994). The existence of those different variants in a particular virus preparation could be explained by mutations or selection, during the replication process in the host system. Those mutations can originate a predominance of certain types of virus variants with surface spikes exhibiting receptor affinity for erythrocytes from specffic origins at different pH values, exhibiting configurations or characteristic types of cell receptors (Markwell et al. 1986, Rogers & D'Souza 1989). However, Muller and Stanley (1944) demonstrated that a minor effect of pH that occurred over the 6.0-8.0 pH range. Domingo et al. (1983) and Steinhauer et al. (1989), during viral genome analysis, could demonstrated that the majority of the RNA virus populations have a high number of sites of pos- sible mutations, which originate heterogenicity of phenotypes in virus preparations. The results from Table show that the hemolylic activity by NDV is a pH-independent process (Lenard & Miller 1981, Junankar & Cherry 1986, Marsh & Pelchen-Matthews 1993). Different strains of NDV exhibited high hemolysis activity at acid pH (5.0), which was too similar between S093 (33.07%) and La Sota (40.72%) samples. However, that B1 sample exhibited low hemolytic activity at that pH value (1.86%). Moreover, table shows that B1 strain of NDV, which is usually weakly hemolytic at acid pH, exhibited a higher hemolytic activity (47.6%) when it was immediately exposed to neutral pH variation, although the pH-independent character of the paramyxoviruscell fusion process is known. This observation could the explained by dissociation between virus particles with better exhibilion of fusogenic sites (F) on virus membranes (Edwards et al. 1983, Haywood 1994, Stegmann 1994). The structure responsible for fusion process developed by structures as influenzavirus hemagglutinin (HA), a fusion peptide, presents high homology. However analysis on receptor binding and fusogenic characters of influenza virus strains shows variability which is explained by low homology of non fusogenic sequences of HA structures. Structures of H3N2 (X-31 strain of A/Aichi/ 68) and H2N2 (A J Japan/305/57) strains of influenza virus exhibited clear differences when analyzed by hemagglutination and hemolysis assays. HA structures of X-31 binds to sialo glycoproteins causing aggregation in the vital membrane. self-aggregation that inhibits those receptor binding (hemagglutination) and fusogenic (hemolysis) processes (Puri et al. 1990). B1 sample could be a HN variant between NDV samples analyzed, showing aggregation of HN structures in the viral membrane at acid pH which did not occur for La Sota and S0-93 strains. A possible explanation for low hemolysis could be that fusogenic process was inhibited by aggregation of HN slruclures, affirming a dependence between HN and the fusogenic process developed by F structures which exhibit high homology at aminoacid level (Hu et al. 1992. Vainionpaa & H)Tpia 1994). Figure 2 shows the receptor binding activity of B1 HN structures which was inhibited at acid pH, showing positivity at higher values of pH. B1 sample that showed the lowest hemagglutination titers at neutral pH was analyzed at large pH range.
Hemolysis percentages exhibited for Bt, S0-93 and LS strains of Newcastle Disease viruses at different pH values at 5.0 - 6.0 pH variation^a
-----------------------------------------------------------
Hemolysis percentage at
Virus ------------------------------------
samples^b pH pH/variation
-----------------------------------------------------------
5.0 5.0 - 6.0
B1 0.00 47.60
S0-93 33.07 ND
LS 40.72 ND
------------------------------------------------------------
a: different pH values were adjusted and pH variations
were obtained using Sorensen phosphate buffer (Pintoet al. 1994), 5M HCl and 1N NaOH, respectively b: the virus samples were used at 5 ug/ml protein concentration ND - not done
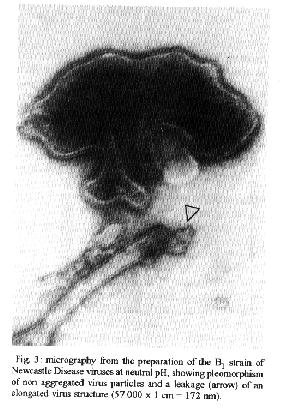
B1 sample already analyzed as above by hemagglutination and hemolysis assays was observed by electron microscopy. The purification process and freezing-thawing cycles developed at neutral pH are shown as important steps to attain disag- gregation and disruption of the pleomorphic virus structure, which induces leakages that are necessary for the liberation of hemoglobin with easy revealation of the virus-cell fusogenic process (Homma et at. 1976). The micrography from a non aggegated preparation of B1 strain of NDV showed a leakage (arrow) that can be observed on an elongated virus particle close to an integral one (Fig. 3). Alterations that were certainly present in all virus preparations submitted at the concentration processes as those evaluated here. ACKNOWLEDGEMENTS To Elizabeth Britto Ferrarezi and Vera Lucia Silva da Cruz for office facilities. To Dr Ary Moreira de Souza (Dept. de Virologia, Empresa Brasileira de Agropecuaria, R J) for helpful discussions. REFERENCES Bradford MM 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye-binding. Anal Biochem 72: 248-254. Clark E, Nagler FPO 1943. Haemagglutination by viruses. The range of susceptible cells with special reference to agglutination by vaccinia virus. Aust J. Exp Biol Med Sci 21: 103-106. ClaveIi LA, Bratl MA 1972. Hemolyfic interaction of Newcastle Disease virus and chicken erythrocytes.II. Determining factors. Appl Immunol 23: 461-470. Couceiro ESS, Coueeiro .INSS, Cabral MC 1994. Myxo and paramyxovirus surface structures: Antigenie and functional studies. Rev Microbiol 25: 24-30. Coueeiro JNSS, Paulson JC, Baum LG 1993. Influenza virus strains selectively recognize sialyloligosaccharides on human respiratory epithelium: the role of the host cell in selection of hemagglutinin receptor specificity. Virus Res 29:155-165. Chu CM 1948. Agglutination of red blood cells of different animal species by influenza and Newcastle Disease viruses. J Hyg 46: 239-246. Domingo E, Martinez-Salas E, Sobrinho F. La Tone JC, Augustin P, Ortin J, Lopez-Galindez C, Perez-Brena P, Villanueva N, Najera R, Scott V, Steinhauer D, Polo N Holland J 1983. The quasiespecies (extremely heterogeneous). Nature of vital RNA genome populations: biological relevance. A Review. Gene 40: 1-8. Edwards J, Mann E, Brown DT 1983. Conformational changes in Sindbis virus envelope proteins accompanying exposure to low pH. J Virol 45: 1090-1097. Fonseca MEF, Frimer N. Mendonga REA, Couceiro J'NSS, Machado RD 1984. A combined staining technique developed for virus particle observation in the electron microscope. Rev Bras Biol 44: 3740. Haywood AM 1994. Virus receptors: Binding adhesion strengthening and changes in viral structure. J Virol 68: 1-5. Homma M, Shimizu K, Shimizu YK, Ishida N 1976. On the study of Sendai virus hemolysis. I. Complete Sendai virus lacking hemolytic activity. Virology 71: 41-42. Hu X, Ray R, Compans RW 1992. Functional interactions between the fusion protein and hemagglutinin-neuraminidase of human Parainfluenza viruses. J Virol 60: 1528-1534. Junankar PR, Cherry RT 1986. Temperature and pH dependence of the hemolytic activity of Influenza virus and the rotational mobility of the spike glycoproteins. Biochem Biophys Acta 854:198. Katz JM, Webster RG 1988. Antigenie and structural characterization of multiple subpopulations of H3N2 influenza virus from an individual. Virology 165: 446-456. Lenard J, Miller DK. 1981 pH dependent hemolysis by influenza. Semliki Forest and Sendai Viruses. Virologly 110: 479-482. Markwell MAK, Moss J, Hom BE, Fishman PH, Svenerholm L 1986. Expression of gangliosides as receptor at the cell surface controls infection of NCTC 2071 cells by Sendai virus. Virology, 155: 356 - 364. Marsh F, Helenius A 1989 Virus entry into animal cells. Adv Virus Res 36: 107-151. Marsh M, Pelchen-Matthews A 1993. Entry of animal viruses into cells. Rev Med Virol 3: 173-185. Martone F, Pacnini F, Bonaduce D, Gatti A 1973. Research into the differentiation of pathogenic and attenuated Newcastle Disease Virus. Atti Soc Ital Sci Vet 27: 618-620. Muller GL, Stanley WM 1944. Quantitative aspects of the red blood cell agglutination test for influenza virus. J Exp Med 79: 185-195. Pinto AMV, Cabral MC, Couceiro JNSS 1994. Hemagglutinating and sialidase activities of subpopulations of influenza viruses. Braz J Med Biol Res 27: 1141-1147.
Phyalla R, Phyalla L, Pekkala P 1988. Host-cell mediated selection of influenza A (H3N2) virus variant subpopulations: lack of association between antigenie and receptor-binding properties. Epidetniol Infect 100:511-522. Pun A, Booy P, Doms RW, White YM, Blumenthal R 1990. Confonnational changes and fusion of influenza virus hemagglutinin of the H2 and H3 subtypes: effects of acid treatment. J Virol 64: 3824-3832. Rogers GN, D'Souza BL 1989. Receptor binding properties of human and animal H1 influenza virus molares. Virology 173: 317-322. Stegmann T 1994. Anchors aweigh. A recent study using a version of the influenza virus hemagglutinin protein in which its membrane anchor was replaced by a lipid tail provides new insights into how proteins can mediate the fusion of membranes. Curr Biol 4: 551-554. Steinhauer DA, Torre JC, Meier E, Holland JJ 1989. Extreme heterogeneity in populations of vesicular stomatitis virus. J Virol 63: 2072-2080. Vainionpaa R, Huyypia T 1994. Biology of parainfluenza viruses. Clin Microbiol Rev 7: 265-275. Winslow NS, Hanson RP, Upton E, Brandly CA 1943. Agglutination of mammalian erythrocytes by Newcastle Disease virus.Aust J Exp Biol Med Sci 21: 174-178.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95102c.gif]Photo images[oc95102c.jpg]Line drawing images[oc95102a.gif] [oc95102b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}